

Cochlear organoids reveal transcriptional programs of postnatal hair cell differentiation from supporting cells
- PMID: 37952154
- PMCID: PMC11007545
- DOI: 10.1016/j.celrep.2023.113421
Cochlear organoids reveal transcriptional programs of postnatal hair cell differentiation from supporting cells
Abstract
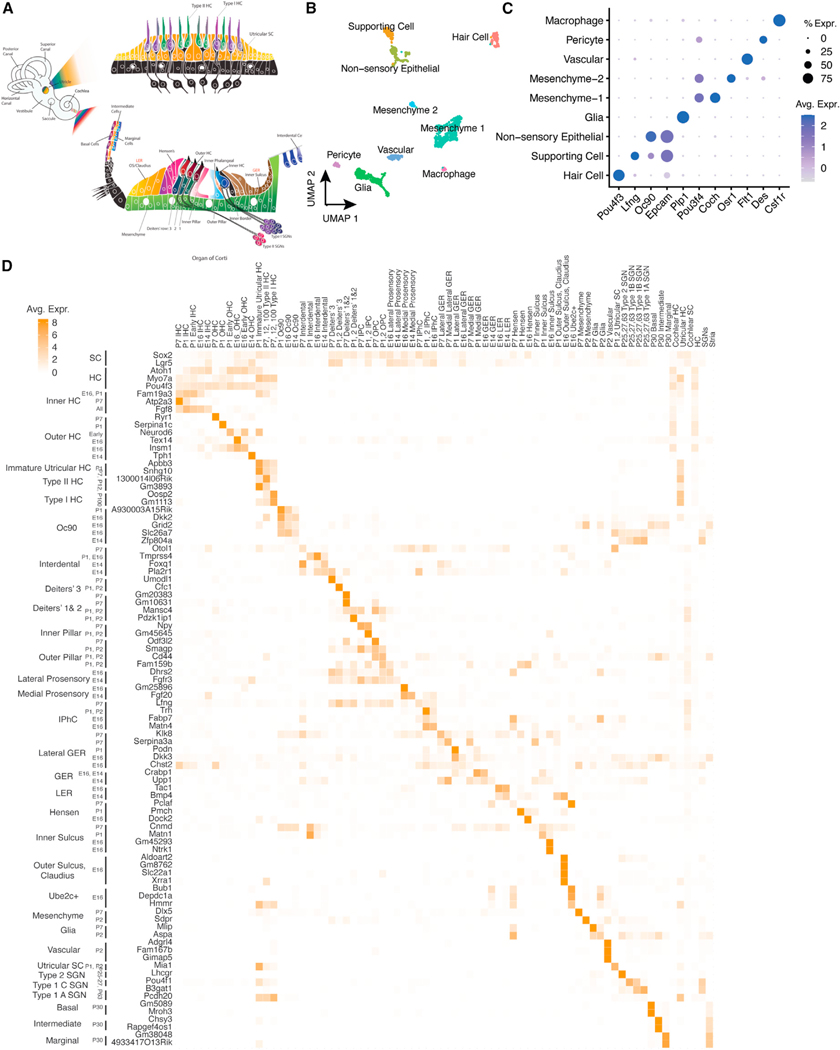
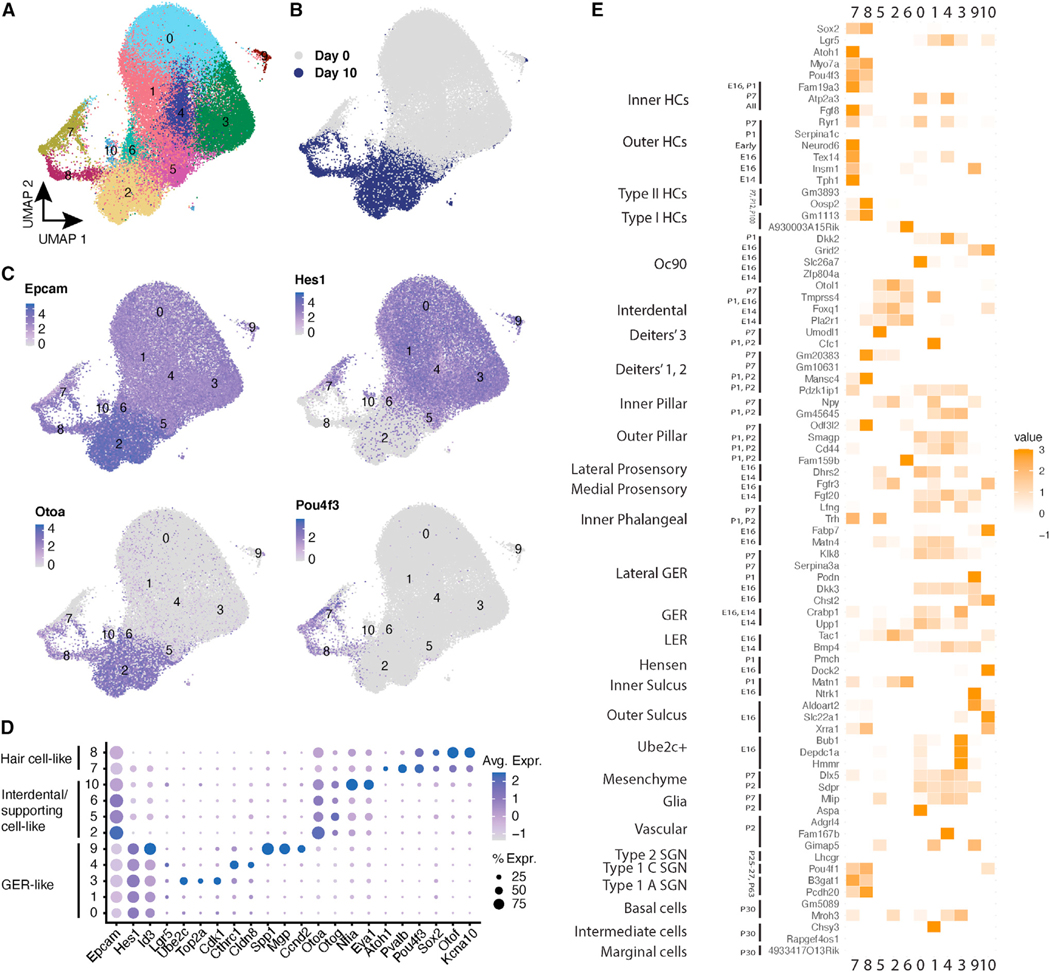
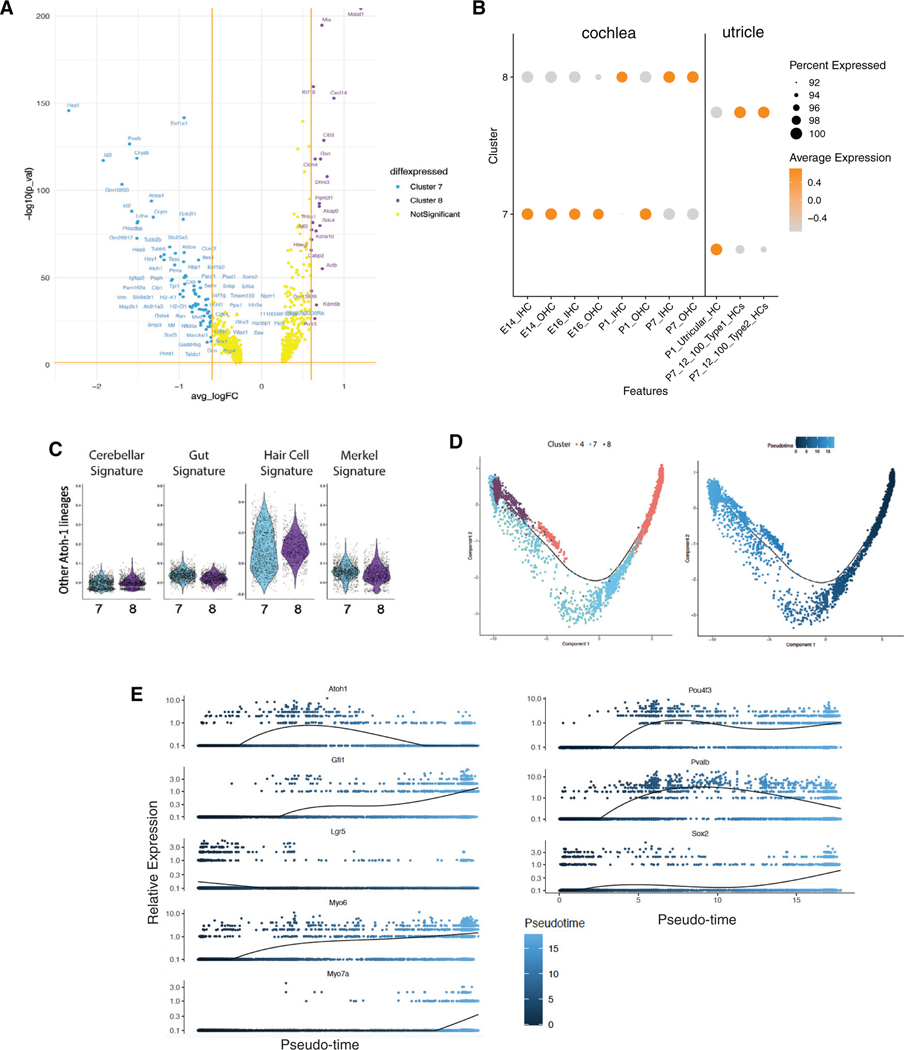
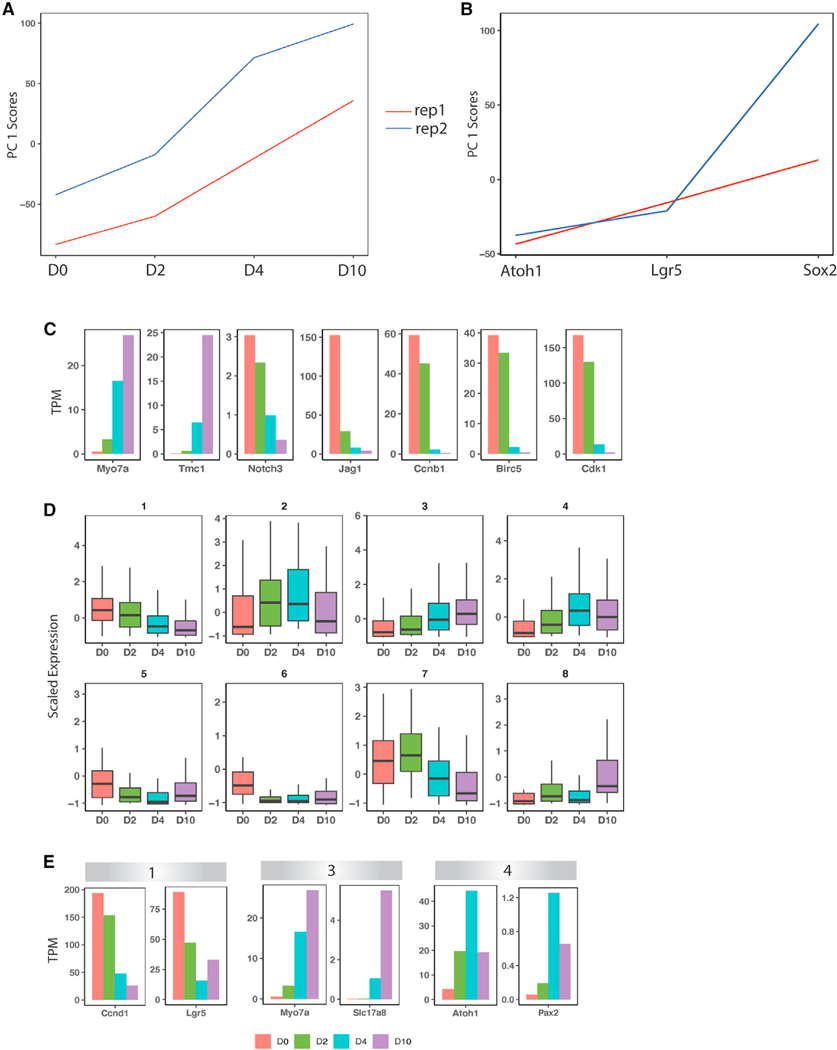
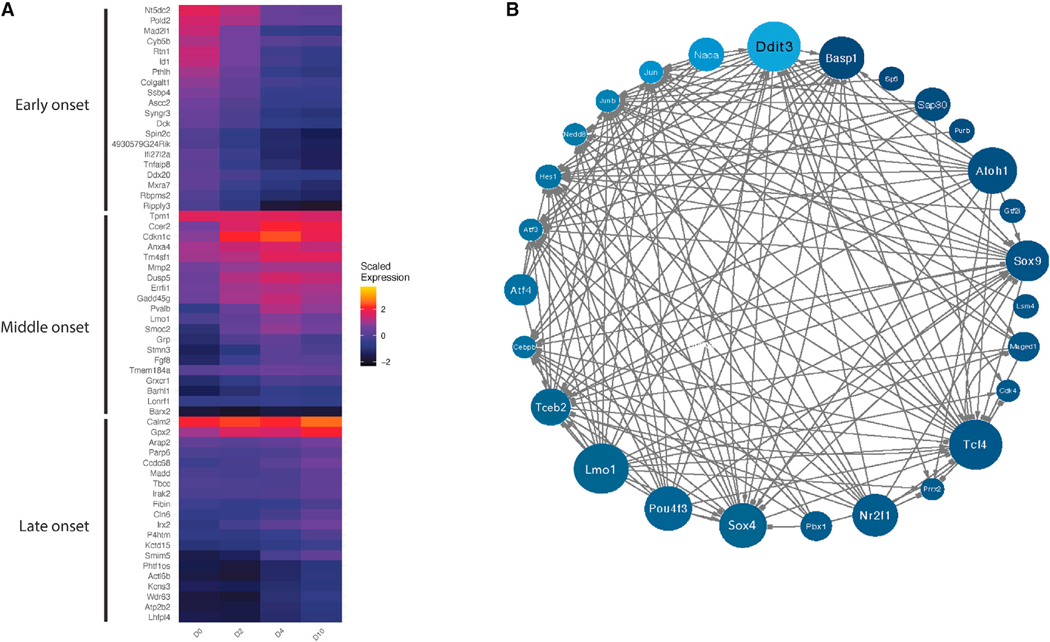
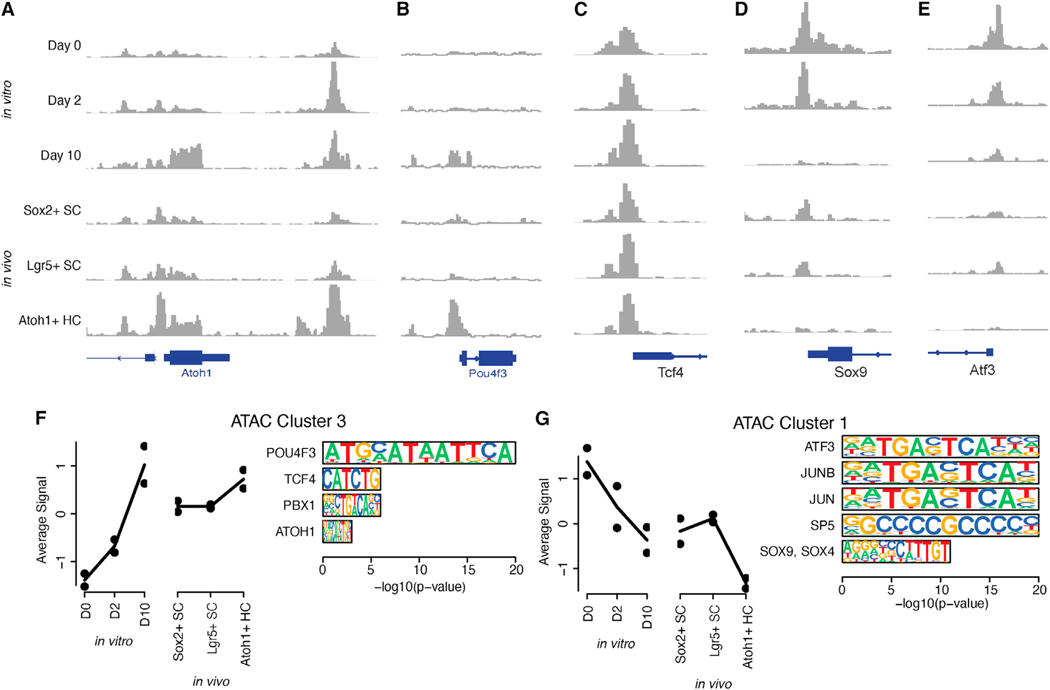
We explore the changes in chromatin accessibility and transcriptional programs for cochlear hair cell differentiation from postmitotic supporting cells using organoids from postnatal cochlea. The organoids contain cells with transcriptional signatures of differentiating vestibular and cochlear hair cells. Construction of trajectories identifies Lgr5+ cells as progenitors for hair cells, and the genomic data reveal gene regulatory networks leading to hair cells. We validate these networks, demonstrating dynamic changes both in expression and predicted binding sites of transcription factors (TFs) during organoid differentiation. We identify known regulators of hair cell development, Atoh1, Pou4f3, and Gfi1, and the analysis predicts the regulatory factors Tcf4, an E-protein and heterodimerization partner of Atoh1, and Ddit3, a CCAAT/enhancer-binding protein (C/EBP) that represses Hes1 and activates transcription of Wnt-signaling-related genes. Deciphering the signals for hair cell regeneration from mammalian cochlear supporting cells reveals candidates for hair cell (HC) regeneration, which is limited in the adult.
Keywords: ATAC sequencing; CP: Stem cell research; cochlea; gene expression analysis; sensory hair cells; single-cell RNA sequencing.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
