

Meta-reinforcement learning via orbitofrontal cortex
- PMID: 37957318
- PMCID: PMC10689244
- DOI: 10.1038/s41593-023-01485-3
Meta-reinforcement learning via orbitofrontal cortex
Erratum in
-
Author Correction: Meta-reinforcement learning via orbitofrontal cortex.Nat Neurosci. 2024 Aug;27(8):1632. doi: 10.1038/s41593-024-01718-z. Nat Neurosci. 2024. PMID: 38956167 Free PMC article. No abstract available.
Abstract
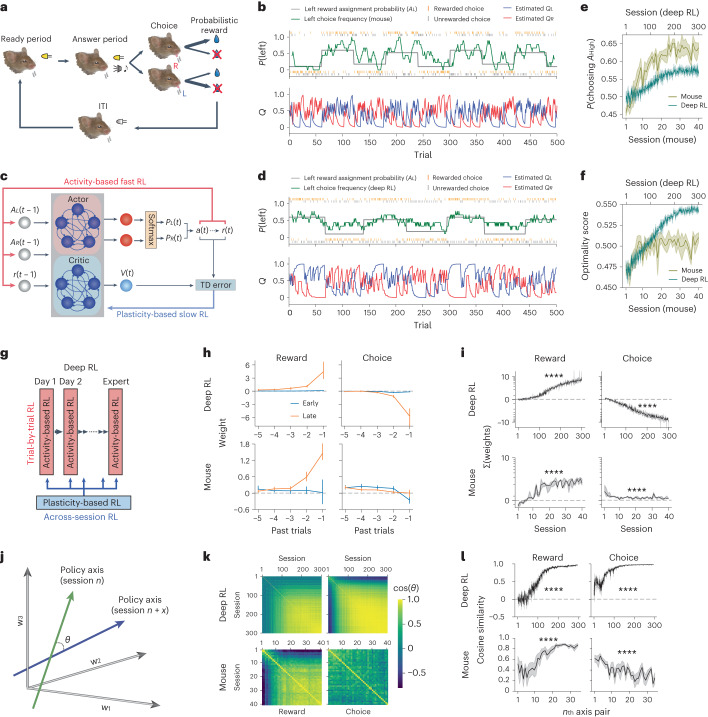
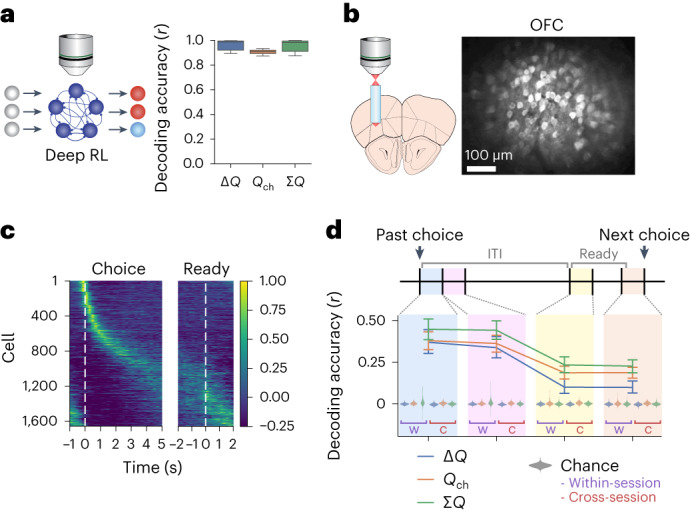
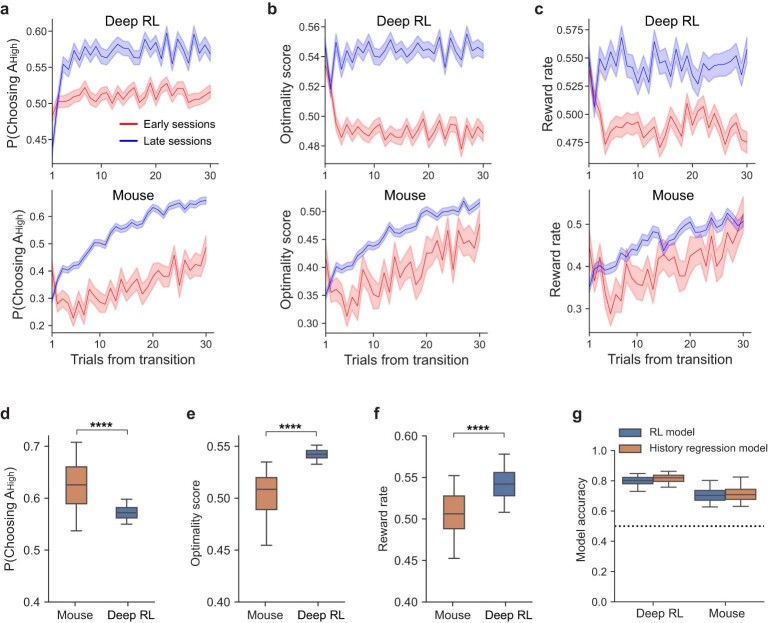
The meta-reinforcement learning (meta-RL) framework, which involves RL over multiple timescales, has been successful in training deep RL models that generalize to new environments. It has been hypothesized that the prefrontal cortex may mediate meta-RL in the brain, but the evidence is scarce. Here we show that the orbitofrontal cortex (OFC) mediates meta-RL. We trained mice and deep RL models on a probabilistic reversal learning task across sessions during which they improved their trial-by-trial RL policy through meta-learning. Ca2+/calmodulin-dependent protein kinase II-dependent synaptic plasticity in OFC was necessary for this meta-learning but not for the within-session trial-by-trial RL in experts. After meta-learning, OFC activity robustly encoded value signals, and OFC inactivation impaired the RL behaviors. Longitudinal tracking of OFC activity revealed that meta-learning gradually shapes population value coding to guide the ongoing behavioral policy. Our results indicate that two distinct RL algorithms with distinct neural mechanisms and timescales coexist in OFC to support adaptive decision-making.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures














References
-
- Hospedales, T., Antoniou, A., Micaelli, P. & Storkey, S. Meta-learning in neural networks: a survey. Preprint at arXiv10.48550/arXiv.2004.05439 (2020). - PubMed
-
- Wang, J. X. et al. Learning to reinforcement learn. Preprint at arXiv10.48550/arXiv.1611.05763 (2016).
-
- Duan, Y. et al. RL2: fast reinforcement learning via slow reinforcement learning. Preprint at arXiv10.48550/arXiv.1611.02779 (2016).
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
