

The molecular basis for cellular function of intrinsically disordered protein regions
- PMID: 37957331
- PMCID: PMC11459374
- DOI: 10.1038/s41580-023-00673-0
The molecular basis for cellular function of intrinsically disordered protein regions
Abstract
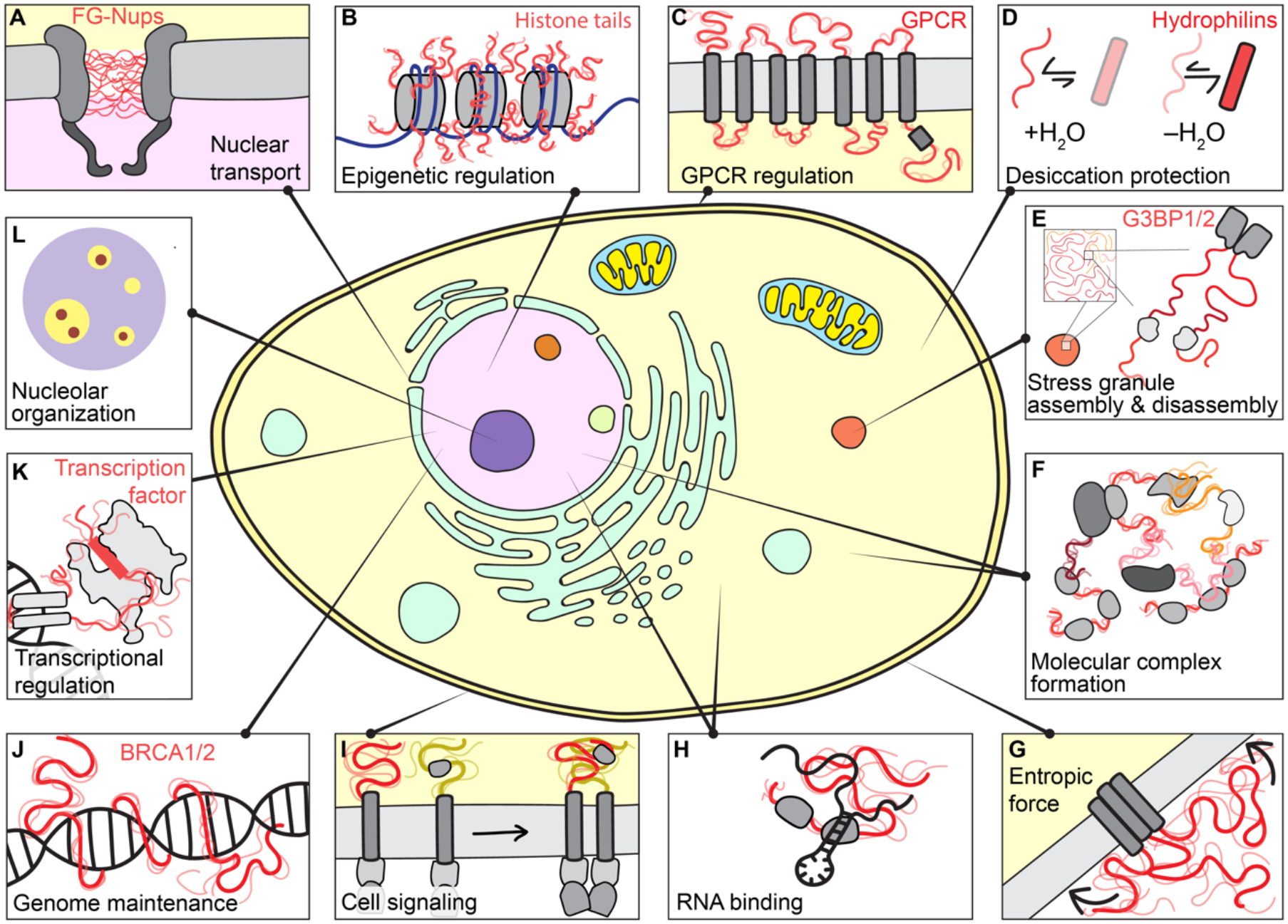
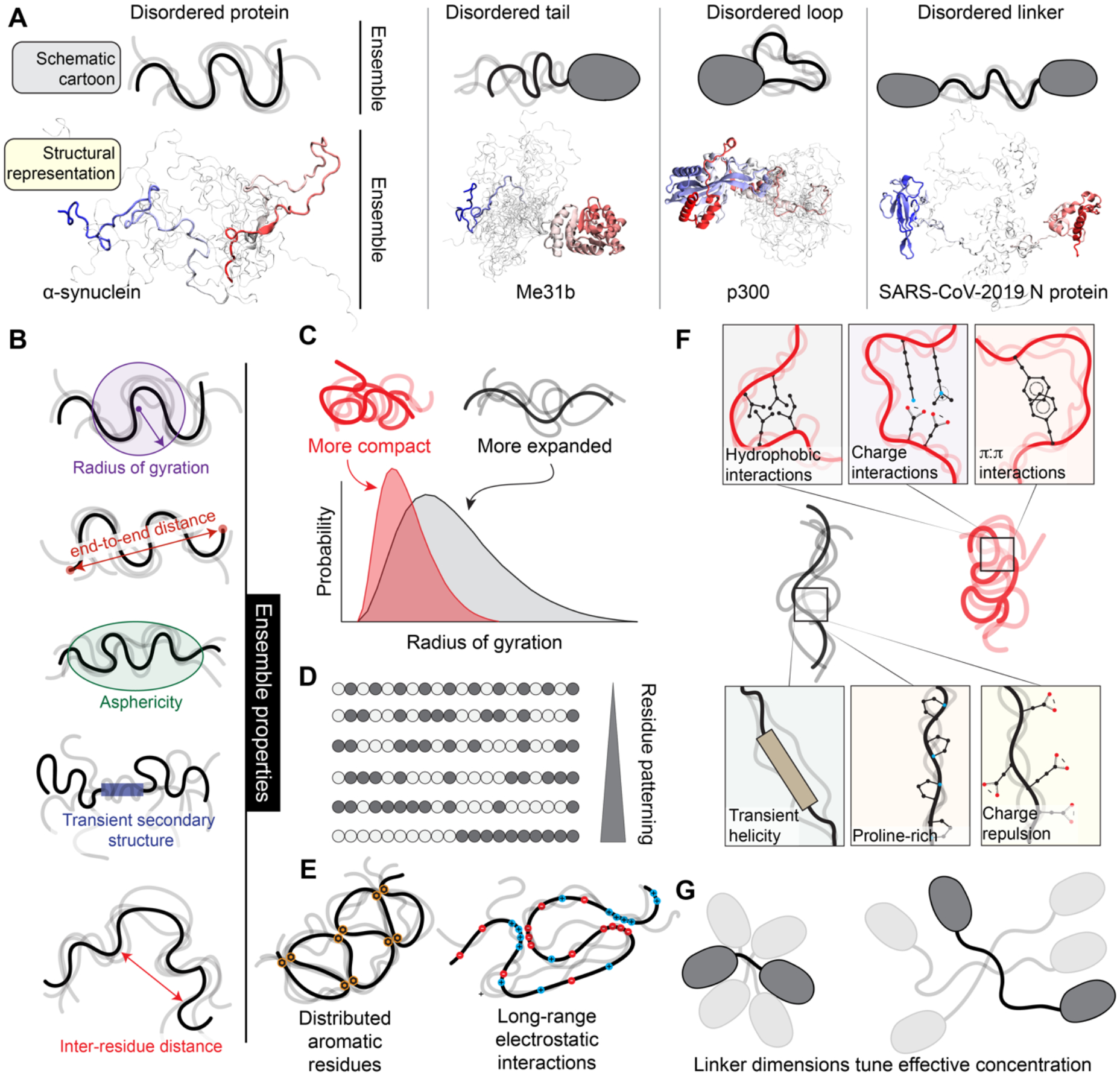
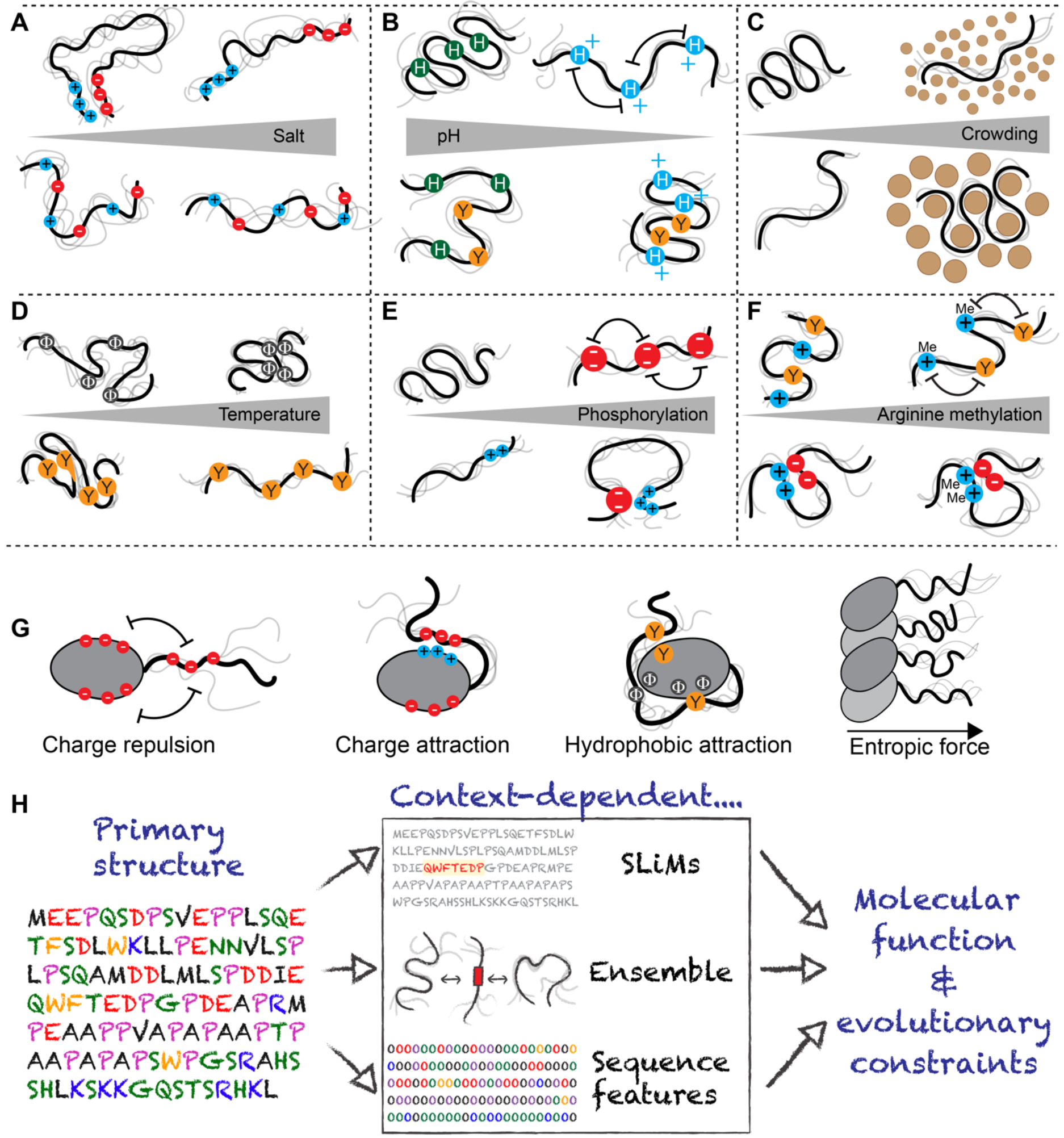
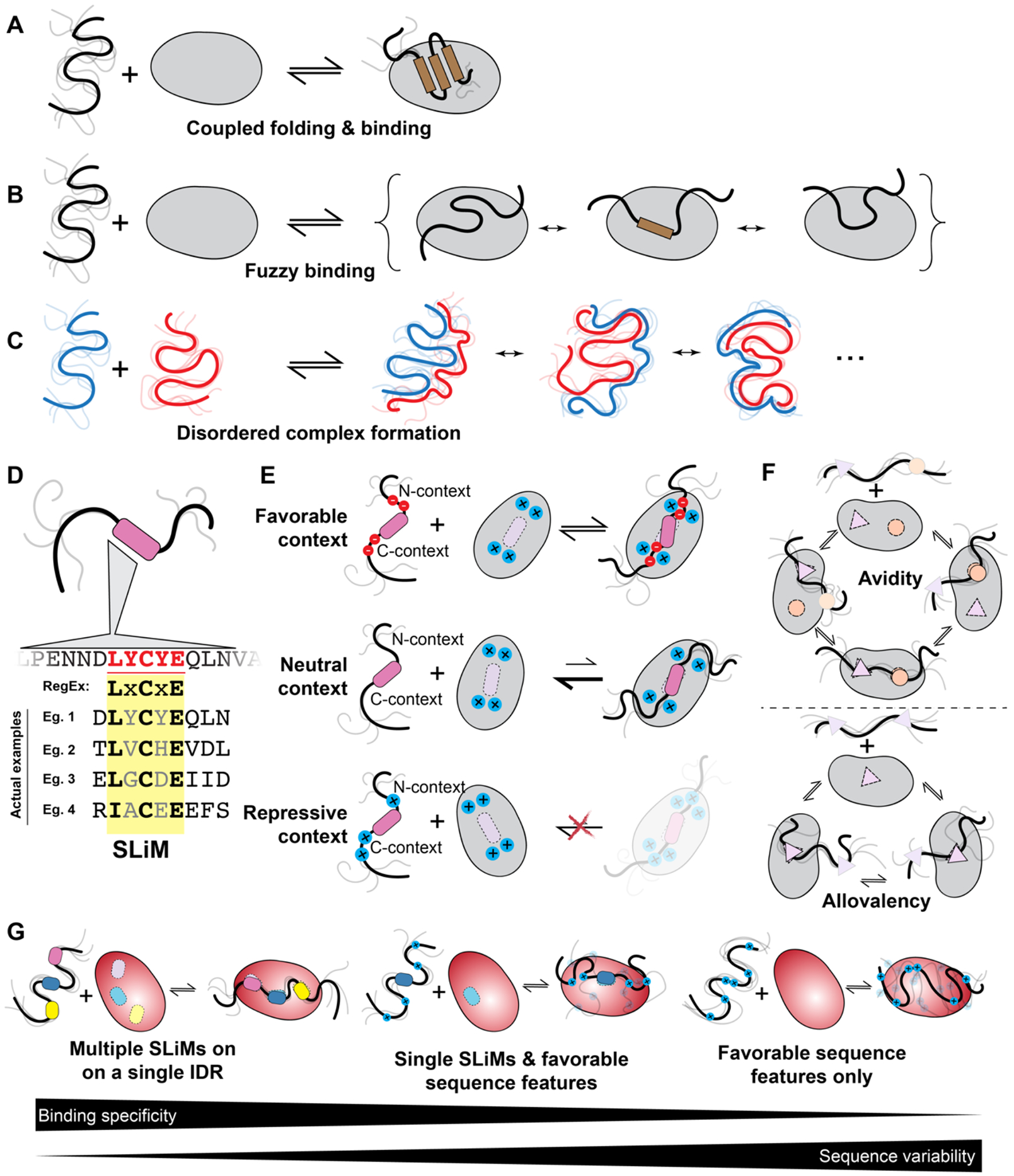
Intrinsically disordered protein regions exist in a collection of dynamic interconverting conformations that lack a stable 3D structure. These regions are structurally heterogeneous, ubiquitous and found across all kingdoms of life. Despite the absence of a defined 3D structure, disordered regions are essential for cellular processes ranging from transcriptional control and cell signalling to subcellular organization. Through their conformational malleability and adaptability, disordered regions extend the repertoire of macromolecular interactions and are readily tunable by their structural and chemical context, making them ideal responders to regulatory cues. Recent work has led to major advances in understanding the link between protein sequence and conformational behaviour in disordered regions, yet the link between sequence and molecular function is less well defined. Here we consider the biochemical and biophysical foundations that underlie how and why disordered regions can engage in productive cellular functions, provide examples of emerging concepts and discuss how protein disorder contributes to intracellular information processing and regulation of cellular function.
© 2023. Springer Nature Limited.
Conflict of interest statement
Competing Interest
A.S.H. is a scientific consultant with Dewpoint Therapeutics and on the Scientific Advisory Board for Prose Foods. All other authors declare no conflicts of interest.
Figures
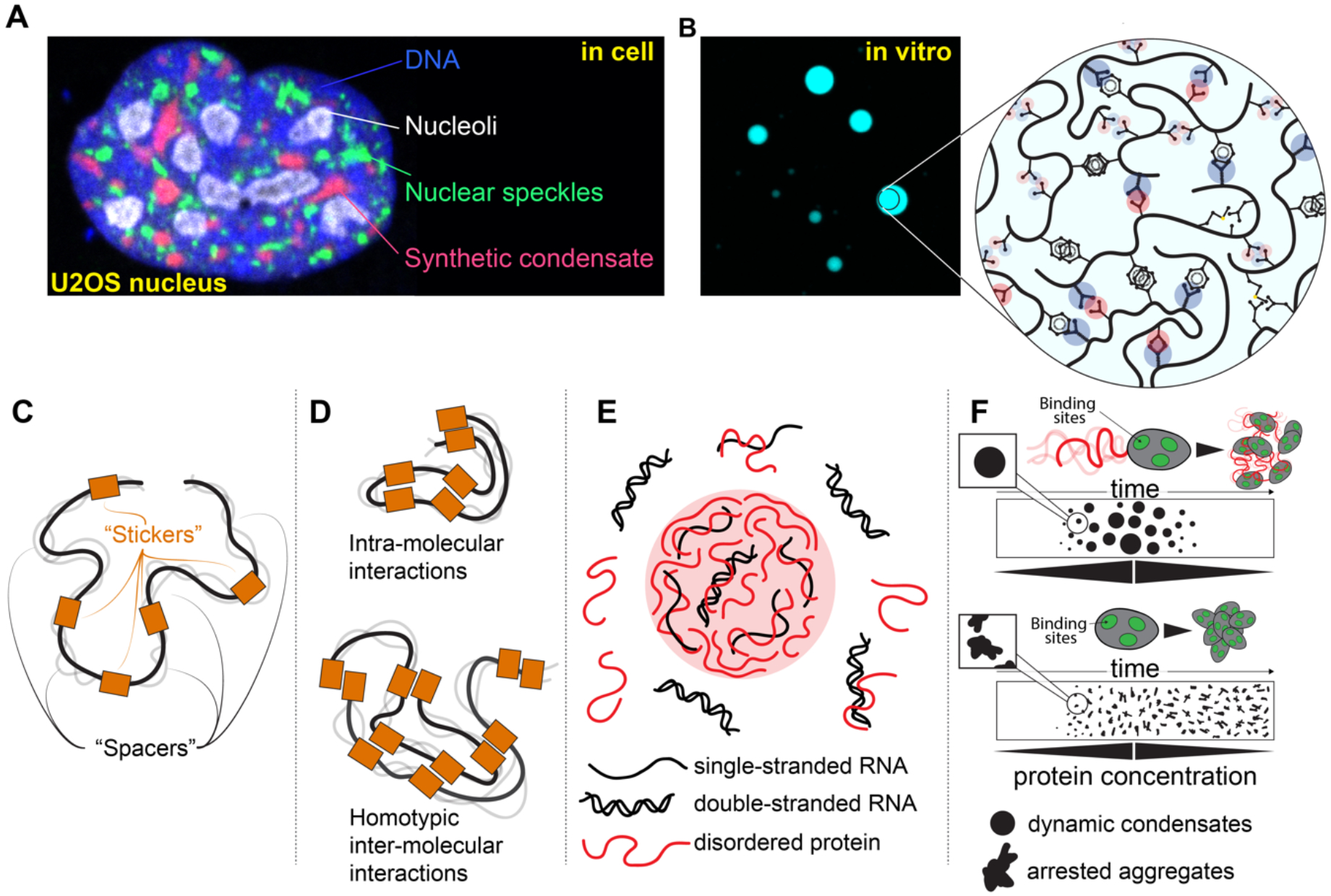
References
Related links
-
- Metapredict disorder predictor: https://metapredict.net/
-
- CAID prediction portal: https://caid.idpcentral.org/submit
-
- Eukaryotic Linear Motif (ELM) resource: http://elm.eu.org/
-
- PLAAC Webserver for identify prion-like domains: http://plaac.wi.mit.edu/
-
- CIDER Webserver for calculating sequence properties: http://pappulab.wustl.edu/CIDER/
References
-
-
Dunker AK et al. Intrinsically disordered protein. J. Mol. Graph. Model 19, 26–59 (2001).
This article (along with Wright & Dyson 1999, Uversky 2001, and Tompa 2002, refs ,, and , respectively) makes the original arguments that IDRs can and do have important roles in cellular function.
-
-
- Wright PE & Dyson HJ Intrinsically unstructured proteins: re-assessing the protein structure-function paradigm. J. Mol. Biol 293, 321–331 (1999). - PubMed
-
- Tompa P. Intrinsically unstructured proteins. Trends Biochem. Sci 27, 527–533 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
