

Transcriptome and Physiological Analysis of Rapeseed Tolerance to Post-Flowering Temperature Increase
- PMID: 37958577
- PMCID: PMC10648292
- DOI: 10.3390/ijms242115593
Transcriptome and Physiological Analysis of Rapeseed Tolerance to Post-Flowering Temperature Increase
Abstract
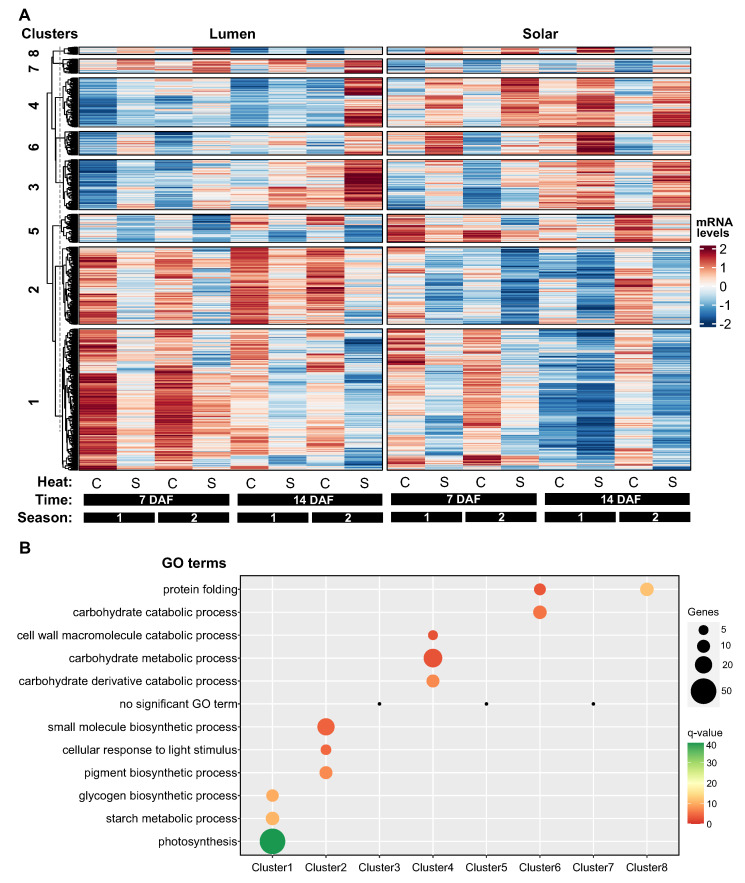
Climate-change-induced temperature fluctuations pose a significant threat to crop production, particularly in the Southern Hemisphere. This study investigates the transcriptome and physiological responses of rapeseed to post-flowering temperature increases, providing valuable insights into the molecular mechanisms underlying rapeseed tolerance to heat stress. Two rapeseed genotypes, Lumen and Solar, were assessed under control and heat stress conditions in field experiments conducted in Valdivia, Chile. Results showed that seed yield and seed number were negatively affected by heat stress, with genotype-specific responses. Lumen exhibited an average of 9.3% seed yield reduction, whereas Solar showed a 28.7% reduction. RNA-seq analysis of siliques and seeds revealed tissue-specific responses to heat stress, with siliques being more sensitive to temperature stress. Hierarchical clustering analysis identified distinct gene clusters reflecting different aspects of heat stress adaptation in siliques, with a role for protein folding in maintaining silique development and seed quality under high-temperature conditions. In seeds, three distinct patterns of heat-responsive gene expression were observed, with genes involved in protein folding and response to heat showing genotype-specific expression. Gene coexpression network analysis revealed major modules for rapeseed yield and quality, as well as the trade-off between seed number and seed weight. Overall, this study contributes to understanding the molecular mechanisms underlying rapeseed tolerance to heat stress and can inform crop improvement strategies targeting yield optimization under changing environmental conditions.
Keywords: Brassica napus; gene coexpression network analysis; heat stress; post-flowering temperature increase; seed number; seed weight; seed yield; transcriptome analysis.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures







References
-
- Cheng W., Sakai H., Yagi K., Hasegawa T. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agric. For. Meteorol. 2009;149:51–58. doi: 10.1016/j.agrformet.2008.07.006. - DOI
-
- Kim H.R., You Y.H. Effects of elevated CO2 concentration and increased temperature on leaf related-physiological responses of Phytolacca insularis (native species) and Phytolacca americana (invasive species) J. Ecol. Environ. 2010;33:195–204. doi: 10.5141/JEFB.2010.33.3.195. - DOI
-
- IPCC . Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press; Cambridge, UK: 2023. - DOI
-
- Rivelli G.M., Long M.E.F., Abeledo L.G., Calderini D.F., Miralles D.J., Rondanini D.P. Assessment of heat stress and cloudiness probabilities in post-flowering of spring wheat and canola in the Southern Cone of South America. Theor. Appl. Climatol. 2021;145:1485–1502. doi: 10.1007/s00704-021-03694-x. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
