

3D reconstruction of murine mitochondria reveals changes in structure during aging linked to the MICOS complex
- PMID: 37960952
- PMCID: PMC10726809
- DOI: 10.1111/acel.14009
3D reconstruction of murine mitochondria reveals changes in structure during aging linked to the MICOS complex
Abstract
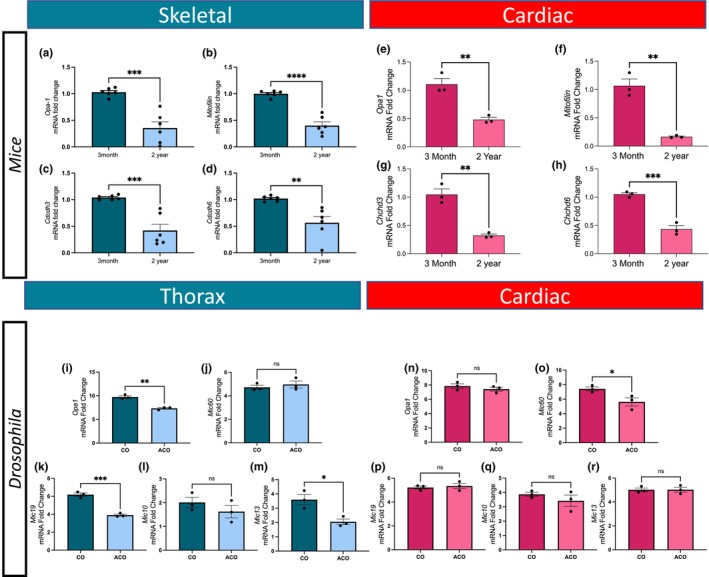
During aging, muscle gradually undergoes sarcopenia, the loss of function associated with loss of mass, strength, endurance, and oxidative capacity. However, the 3D structural alterations of mitochondria associated with aging in skeletal muscle and cardiac tissues are not well described. Although mitochondrial aging is associated with decreased mitochondrial capacity, the genes responsible for the morphological changes in mitochondria during aging are poorly characterized. We measured changes in mitochondrial morphology in aged murine gastrocnemius, soleus, and cardiac tissues using serial block-face scanning electron microscopy and 3D reconstructions. We also used reverse transcriptase-quantitative PCR, transmission electron microscopy quantification, Seahorse analysis, and metabolomics and lipidomics to measure changes in mitochondrial morphology and function after loss of mitochondria contact site and cristae organizing system (MICOS) complex genes, Chchd3, Chchd6, and Mitofilin. We identified significant changes in mitochondrial size in aged murine gastrocnemius, soleus, and cardiac tissues. We found that both age-related loss of the MICOS complex and knockouts of MICOS genes in mice altered mitochondrial morphology. Given the critical role of mitochondria in maintaining cellular metabolism, we characterized the metabolomes and lipidomes of young and aged mouse tissues, which showed profound alterations consistent with changes in membrane integrity, supporting our observations of age-related changes in muscle tissues. We found a relationship between changes in the MICOS complex and aging. Thus, it is important to understand the mechanisms that underlie the tissue-dependent 3D mitochondrial phenotypic changes that occur in aging and the evolutionary conservation of these mechanisms between Drosophila and mammals.
Keywords: Drosophila; 3D morphometry; MICOS; aging; mitochondria; mitochondrial disease; mitochondrion; reconstruction; reticulum; serial block-face SEM; skeletal muscle.
© 2023 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
All authors declare that they have no conflict of interest.
Figures





References
-
- Abrigo, J. , Gonzalez, F. , Aguirre, F. , Tacchi, F. , Gonzalez, A. , Meza, M. P. , Simon, F. , Cabrera, D. , Arrese, M. , Karpen, S. , & Cabello‐Verrugio, C. (2021). Cholic acid and deoxycholic acid induce skeletal muscle atrophy through a mechanism dependent on TGR5 receptor. Journal of Cellular Physiology, 236, 260–272. - PubMed
-
- Adusumilli, R. , & Mallick, P. (2017). Data conversion with ProteoWizard msConvert. In Comai L., Katz J. E., & Mallick P. (Eds.), Proteomics: Methods and protocols (pp. 339–368). Springer. - PubMed
-
- Bannwarth, S. , Ait‐El‐Mkadem, S. , Chaussenot, A. , Genin, E. C. , Lacas‐Gervais, S. , Fragaki, K. , Berg‐Alonso, L. , Kageyama, Y. , Serre, V. , Moore, D. G. , Verschueren, A. , Rouzier, C. , Le Ber, I. , Augé, G. , Cochaud, C. , Lespinasse, F. , N'Guyen, K. , de Septenville, A. , Brice, A. , … Paquis‐Flucklinger, V. (2014). A mitochondrial origin for frontotemporal dementia and amyotrophic lateral sclerosis through CHCHD10 involvement. Brain, 137, 2329–2345. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- UL1 TR000445/TR/NCATS NIH HHS/United States
- DK101003/NH/NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- R01HL144941/NH/NIH HHS/United States
- R03HL155041/NH/NIH HHS/United States
- T32 DK101003/DK/NIDDK NIH HHS/United States
- R01 HL144941/HL/NHLBI NIH HHS/United States
- R01HL147818/NH/NIH HHS/United States
- UL1 RR024975/RR/NCRR NIH HHS/United States
- Grant 2 UL1 TR000445-06/RR/NCRR NIH HHS/United States
- RR024975-01/RR/NCRR NIH HHS/United States
- 5R25HL106365-12/NH/NIH HHS/United States
- R25 NS117367/NS/NINDS NIH HHS/United States
- R25 HL106365/HL/NHLBI NIH HHS/United States
- I01 BX005352/BX/BLRD VA/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- K01AG062757/NH/NIH HHS/United States
- T32 DK007563/DK/NIDDK NIH HHS/United States
- GT15655/HHMI/Howard Hughes Medical Institute/United States
- HD090061/NH/NIH HHS/United States
- DK007563/NH/NIH HHS/United States
LinkOut - more resources
Full Text Sources
