

This is a preprint.
Long-term in vivo three-photon imaging reveals region-specific differences in healthy and regenerative oligodendrogenesis
- PMID: 37961298
- PMCID: PMC10634963
- DOI: 10.1101/2023.10.29.564636
Long-term in vivo three-photon imaging reveals region-specific differences in healthy and regenerative oligodendrogenesis
Update in
-
Long-term in vivo three-photon imaging reveals region-specific differences in healthy and regenerative oligodendrogenesis.Nat Neurosci. 2024 May;27(5):846-861. doi: 10.1038/s41593-024-01613-7. Epub 2024 Mar 27. Nat Neurosci. 2024. PMID: 38539013 Free PMC article.
Abstract
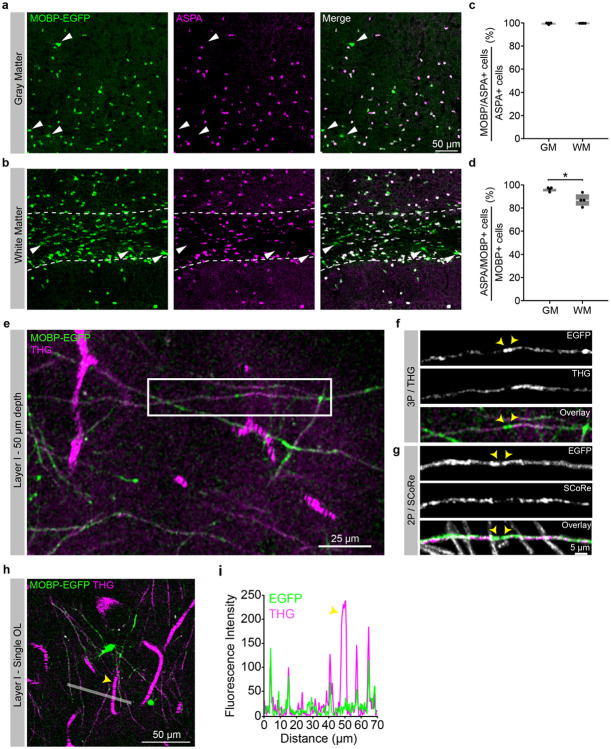
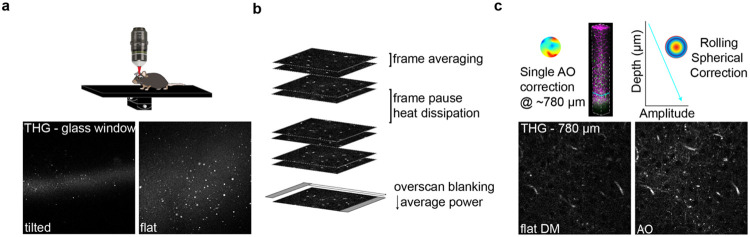
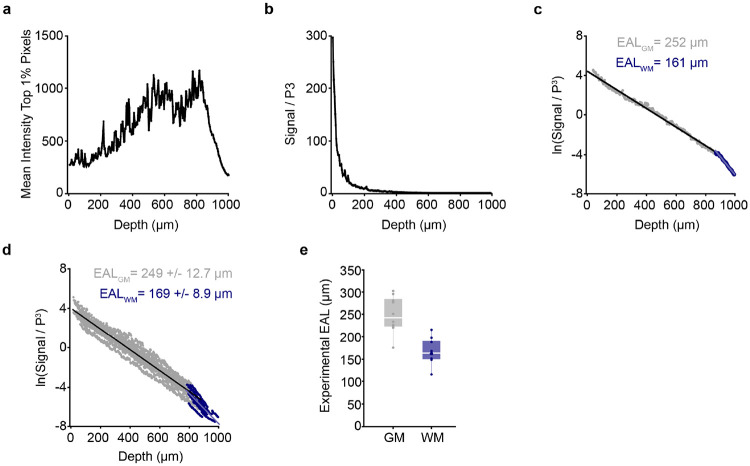
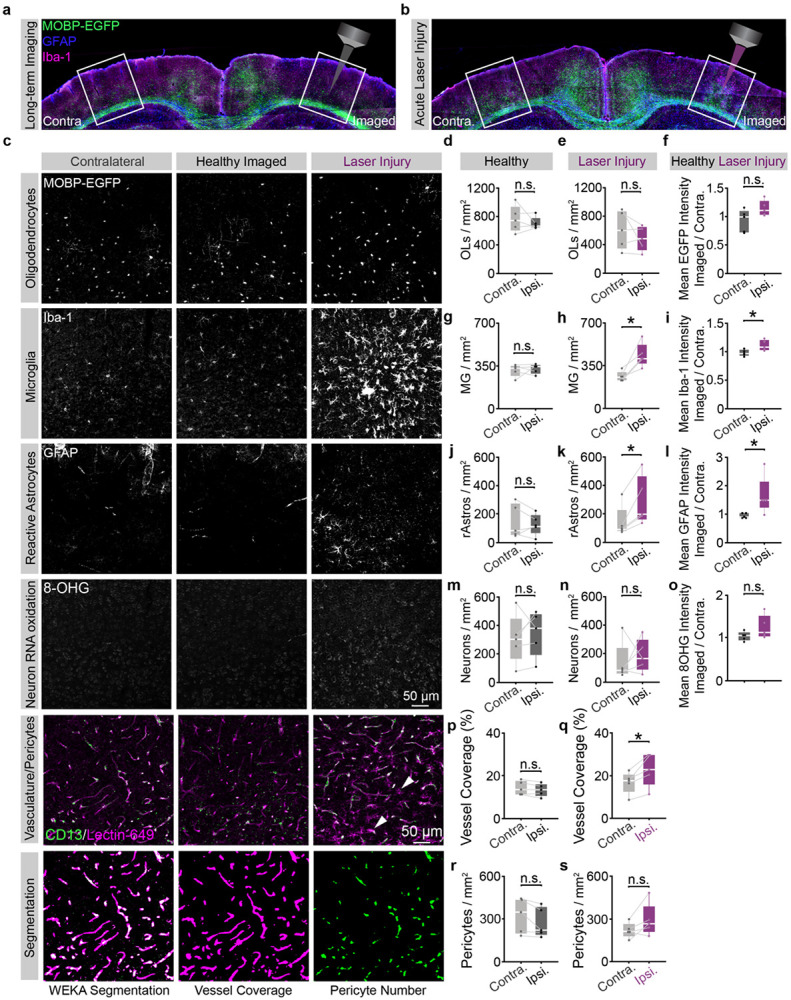
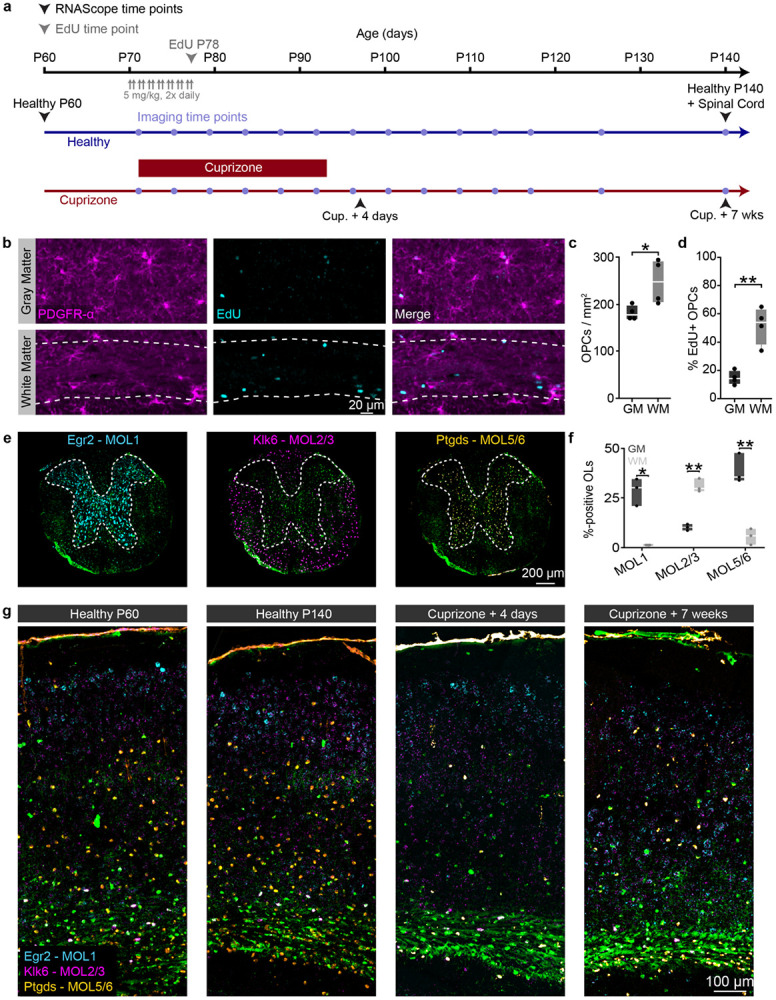
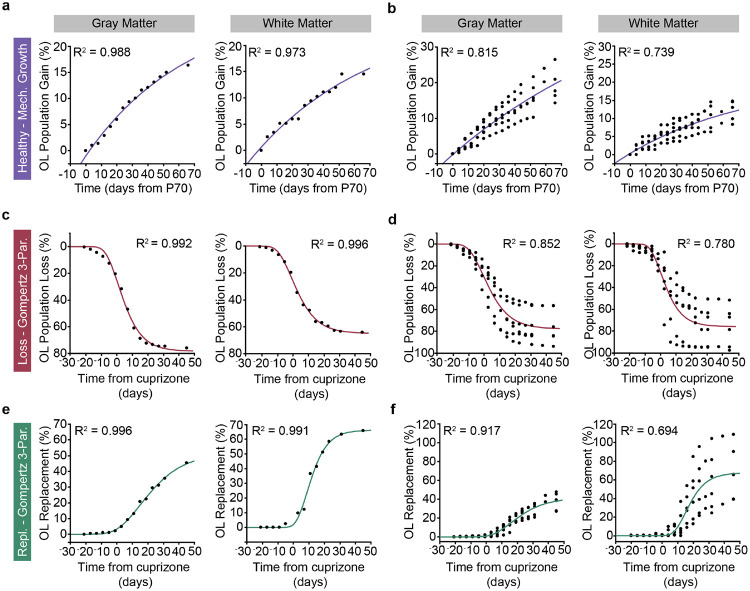
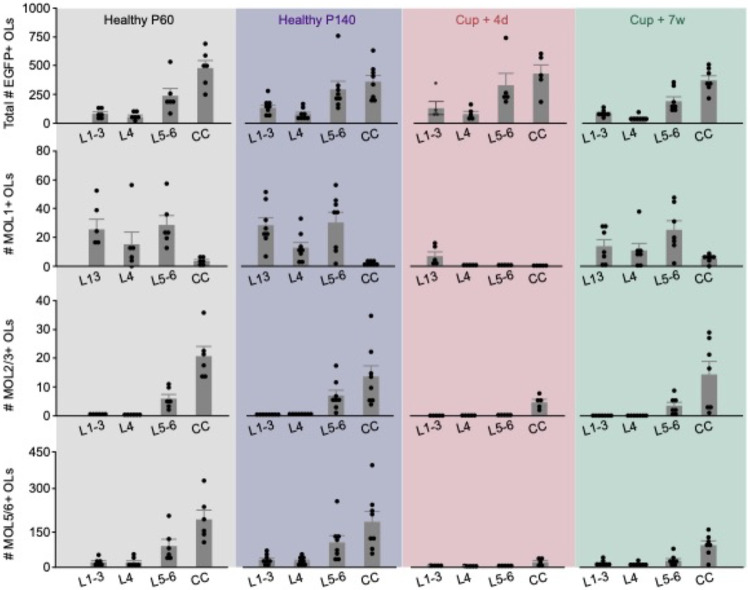
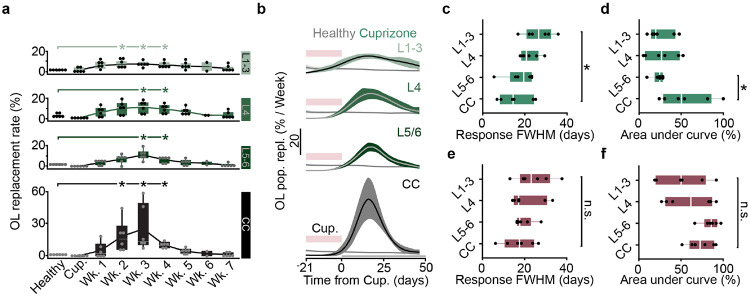
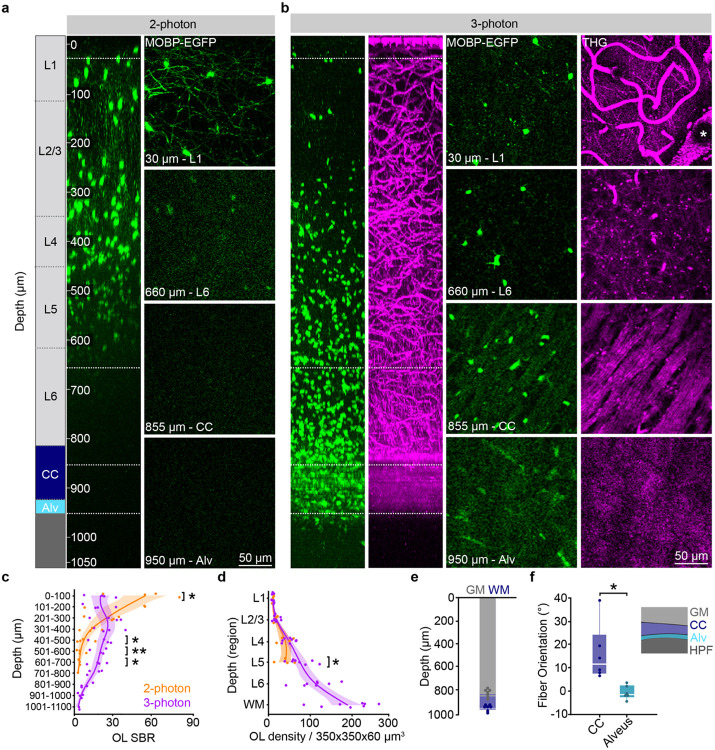
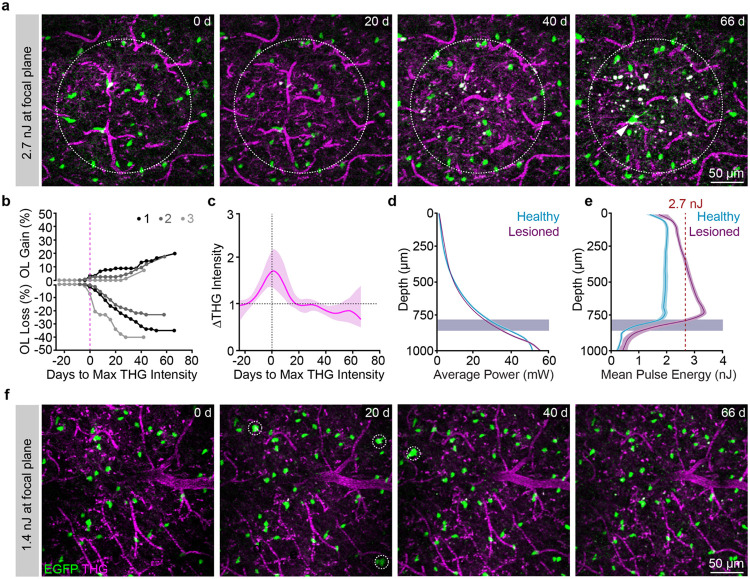
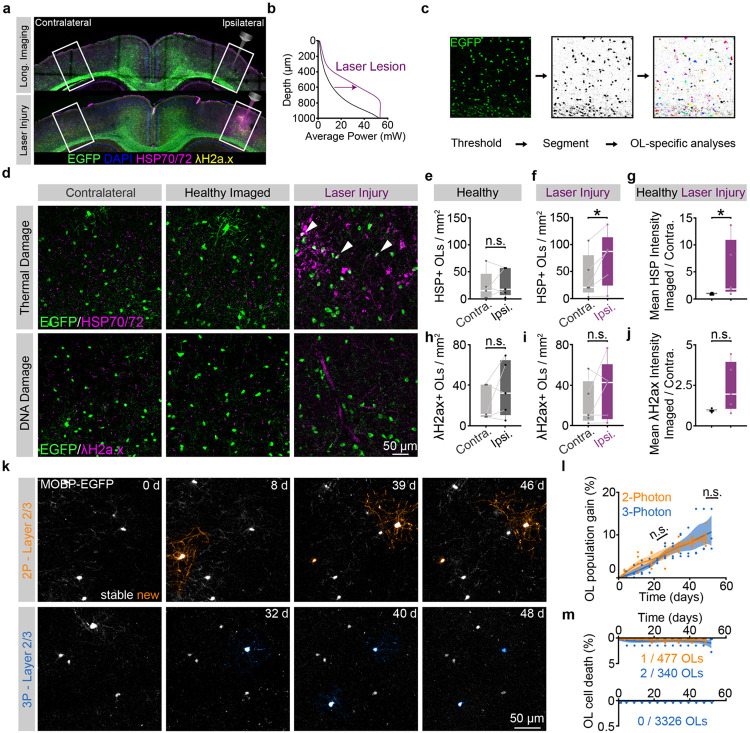
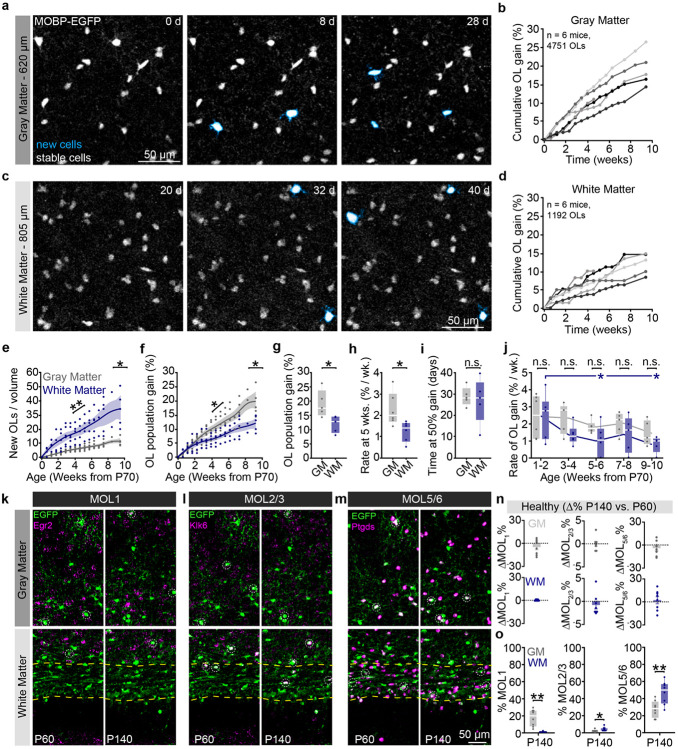
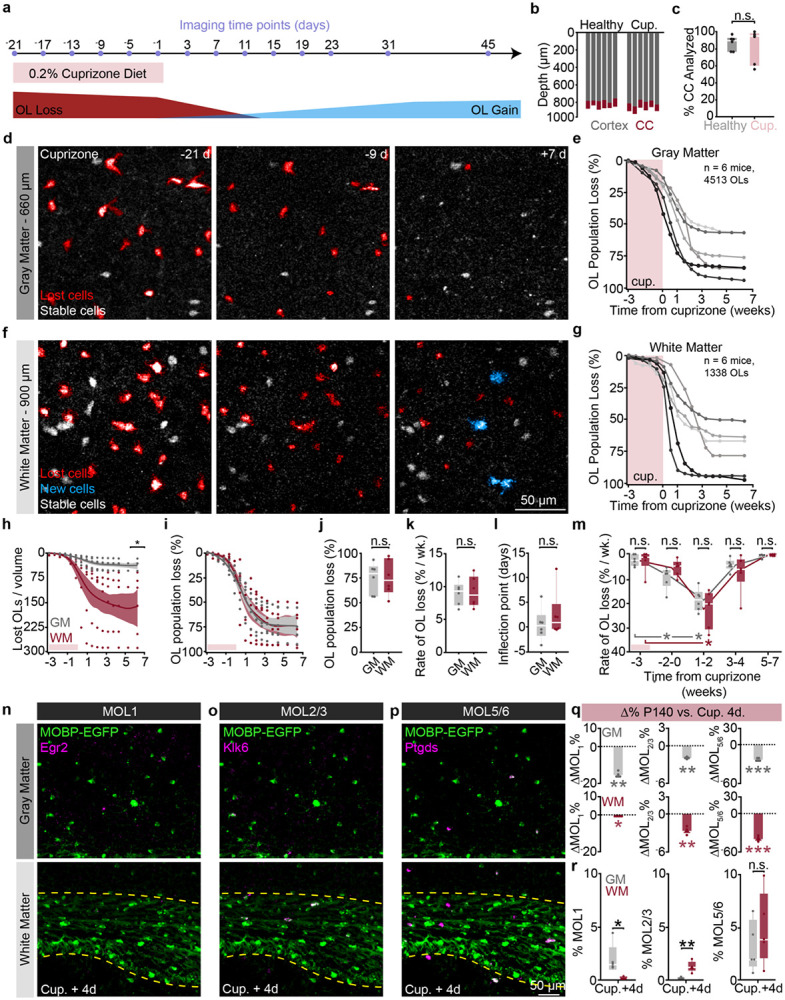
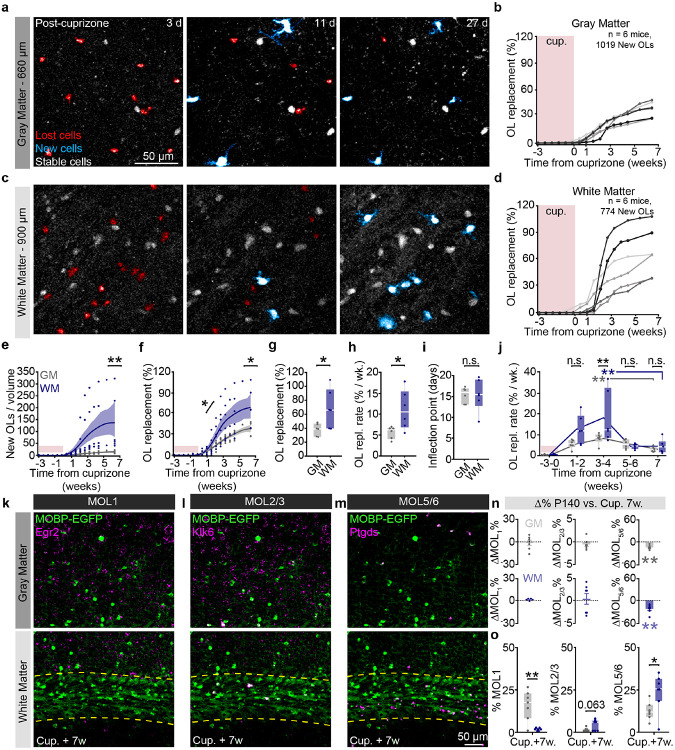
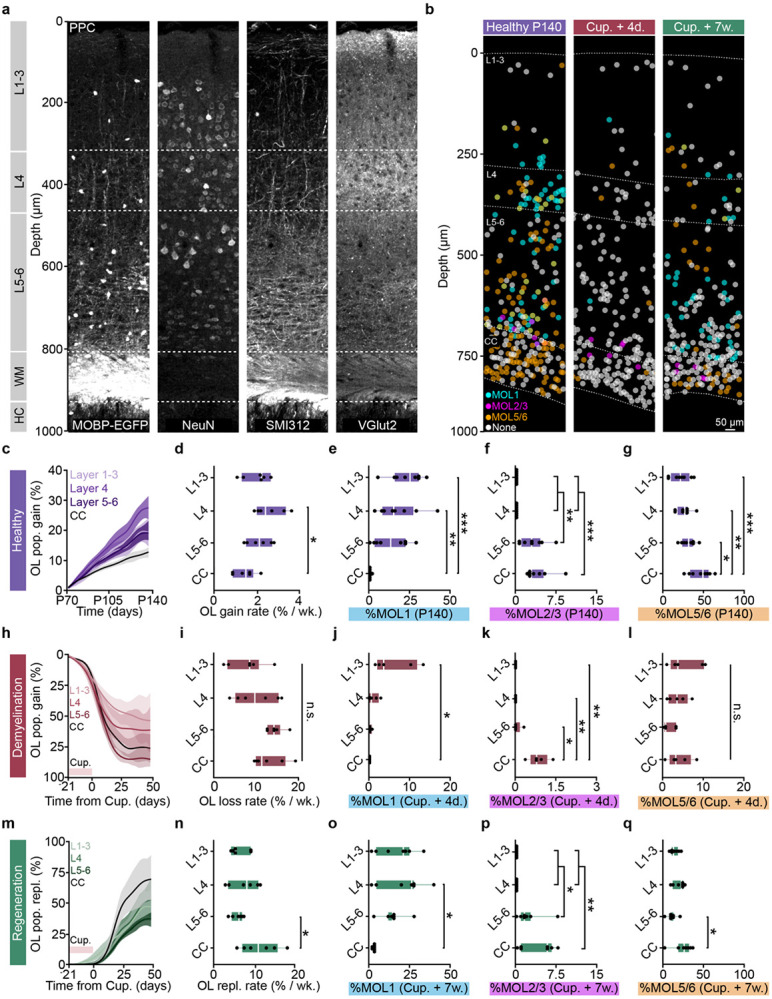
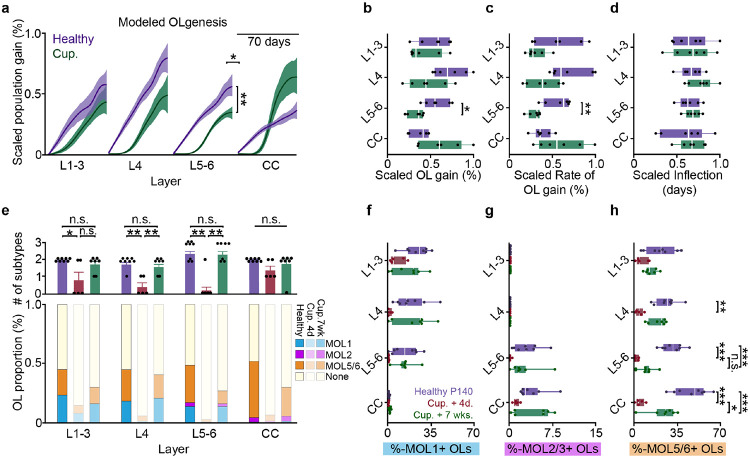
The generation of new myelin-forming oligodendrocytes in the adult CNS is critical for cognitive function and regeneration following injury. Oligodendrogenesis varies between gray and white matter regions suggesting that local cues drive regional differences in myelination and the capacity for regeneration. Yet, the determination of regional variability in oligodendrocyte cell behavior is limited by the inability to monitor the dynamics of oligodendrocytes and their transcriptional subpopulations in white matter of the living brain. Here, we harnessed the superior imaging depth of three-photon microscopy to permit long-term, longitudinal in vivo three-photon imaging of an entire cortical column and underlying subcortical white matter without cellular damage or reactivity. Using this approach, we found that the white matter generated substantially more new oligodendrocytes per volume compared to the gray matter, yet the rate of population growth was proportionally higher in the gray matter. Following demyelination, the white matter had an enhanced population growth that resulted in higher oligodendrocyte replacement compared to the gray matter. Finally, deep cortical layers had pronounced deficits in regenerative oligodendrogenesis and restoration of the MOL5/6-positive oligodendrocyte subpopulation following demyelinating injury. Together, our findings demonstrate that regional microenvironments regulate oligodendrocyte population dynamics and heterogeneity in the healthy and diseased brain.
Figures


References
-
- Flechsig Of Leipsic P. DEVELOPMENTAL (MYELOGENETIC) LOCALISATION OF THE CEREBRAL CORTEX IN THE HUMAN SUBJECT. The Lancet 158, 1027–1030 (1901).
-
- Viganò F., Möbius W., Götz M. & Dimou L. Transplantation reveals regional differences in oligodendrocyte differentiation in the adult brain. Nat Neurosci 16, 1370–1372 (2013). - PubMed
Methods References
-
- Squier J. A. & Müller M. Third-harmonic generation imaging of laser-induced breakdown in glass. Appl. Opt., AO 38, 5789–5794 (1999). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources