

This is a preprint.
Coordinated changes in a cortical circuit sculpt effects of novelty on neural dynamics
- PMID: 37961331
- PMCID: PMC10634721
- DOI: 10.1101/2023.10.21.563440
Coordinated changes in a cortical circuit sculpt effects of novelty on neural dynamics
Update in
-
Coordinated changes in a cortical circuit sculpt effects of novelty on neural dynamics.Cell Rep. 2024 Sep 24;43(9):114763. doi: 10.1016/j.celrep.2024.114763. Epub 2024 Sep 16. Cell Rep. 2024. PMID: 39288028 Free PMC article.
Abstract
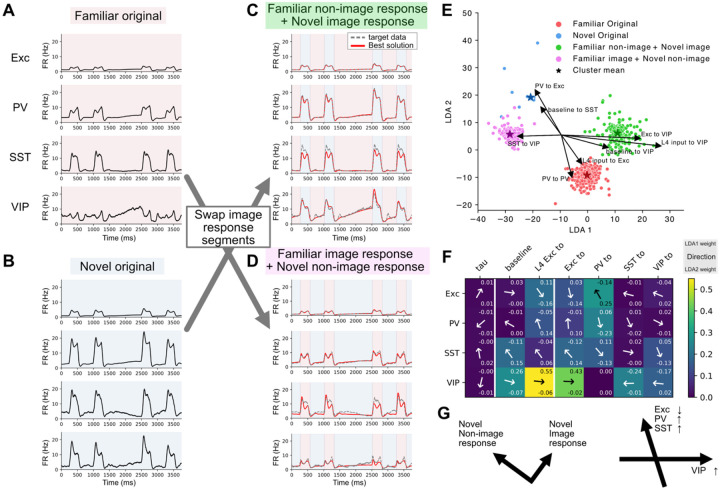
Recent studies have found dramatic cell-type specific responses to stimulus novelty, highlighting the importance of analyzing the cortical circuitry at the cell-type specific level of granularity to understand brain function. Although initial work classified and characterized activity for each cell type, the specific alterations in cortical circuitry-particularly when multiple novelty effects interact-remain unclear. To address this gap, we employed a large-scale public dataset of electrophysiological recordings in the visual cortex of awake, behaving mice using Neuropixels probes and designed population network models to investigate the observed changes in neural dynamics in response to a combination of distinct forms of novelty. The model parameters were rigorously constrained by publicly available structural datasets, including multi-patch synaptic physiology and electron microscopy data. Our systematic optimization approach identified tens of thousands of model parameter sets that replicate the observed neural activity. Analysis of these solutions revealed generally weaker connections under novel stimuli, as well as a shift in the balance e between SST and VIP populations. Along with this, PV and SST populations experienced overall more excitatory influences compared to excitatory and VIP populations. Our results also highlight the role of VIP neurons in multiple aspects of visual stimulus processing and altering gain and saturation dynamics under novel conditions. In sum, our findings provide a systematic characterization of how the cortical circuit adapts to stimulus novelty by combining multiple rich public datasets.
Figures







References
-
- Aitken K., Campagnola L., Garrett M., Olsen S., & Mihalas S. (2023). Familiarity modulated synapses model visual cortical circuit novelty responses [Preprint]. bioRxiv. 10.1101/2023.08.16.553635 - DOI
-
- Billeh Y. N., Cai B., Gratiy S. L., Dai K., Iyer R., Gouwens N. W., Abbasi-Asl R., Jia X., Siegle J. H., Olsen S. R., Koch C., Mihalas S., & Arkhipov A. (2020). Systematic Integration of Structural and Functional Data into Multi-scale Models of Mouse Primary Visual Cortex. Neuron, 106(3), 388–403.e18. 10.1016/j.neuron.2020.01.040 - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous