

This is a preprint.
Rationally-defined microbial consortia suppress multidrug-resistant proinflammatory Enterobacteriaceae via ecological control
- PMID: 37961431
- PMCID: PMC10635318
- DOI: 10.21203/rs.3.rs-3462622/v1
Rationally-defined microbial consortia suppress multidrug-resistant proinflammatory Enterobacteriaceae via ecological control
Update in
-
Commensal consortia decolonize Enterobacteriaceae via ecological control.Nature. 2024 Sep;633(8031):878-886. doi: 10.1038/s41586-024-07960-6. Epub 2024 Sep 18. Nature. 2024. PMID: 39294375 Free PMC article.
Abstract
Persistent colonization and outgrowth of pathogenic organisms in the intestine may occur due to long-term antibiotic usage or inflammatory conditions, which perpetuate dysregulated immunity and tissue damage1,2. Gram-negative Enterobacteriaceae gut pathobionts are particularly recalcitrant to conventional antibiotic treatment3,4, though an emerging body of evidence suggests that manipulation of the commensal microbiota may be a practical alternative therapeutic strategy5-7. In this study, we rationally isolated and down-selected commensal bacterial consortia from healthy human stool samples capable of strongly and specifically suppressing intestinal Enterobacteriaceae. One of the elaborated consortia, consisting of 18 commensal strains, effectively controlled ecological niches by regulating gluconate availability, thereby reestablishing colonization resistance and alleviating antibiotic-resistant Klebsiella-driven intestinal inflammation in mice. Harnessing these microbial activities in the form of live bacterial therapeutics may represent a promising solution to combat the growing threat of proinflammatory, antimicrobial-resistant bacterial infection.
Conflict of interest statement
Competing interests K.H. is a scientific advisory board member of Vedanta Biosciences and 4BIO CAPITAL. Y.A., M.U., K.Ama., and Y.K. are employees of JSR corporation. R.J.X. is co-founder of Jnana Therapeutics and Celsius Therapeutics, scientific advisory board member at Nestlé, and board director at MoonLake Immunotherapeutics. J.M.N, and B.O. are employees of Vedanta Biosciences. S.C. was an employee of Vedanta Biosciences at the time of her contributions. All other authors declare no competing interests.
Figures
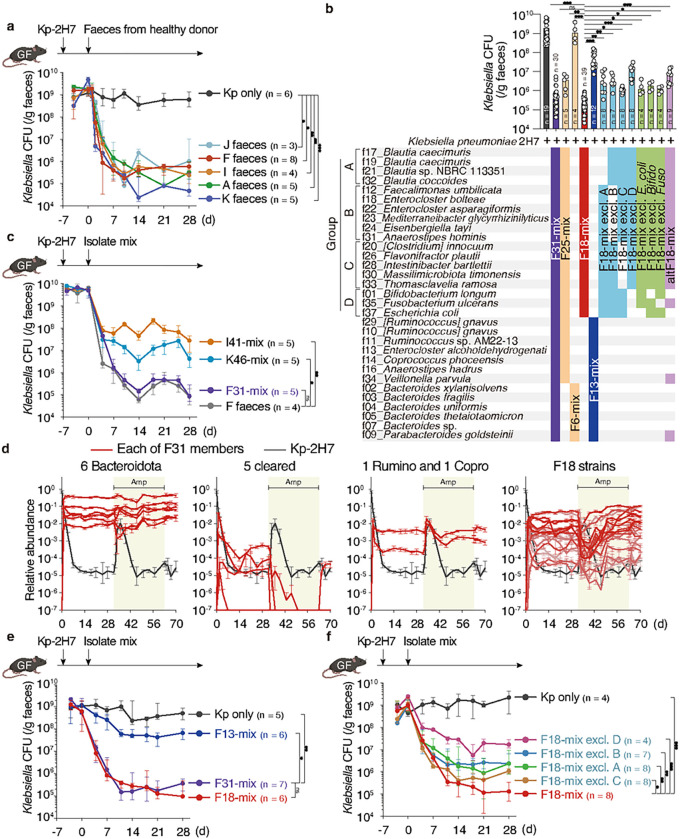
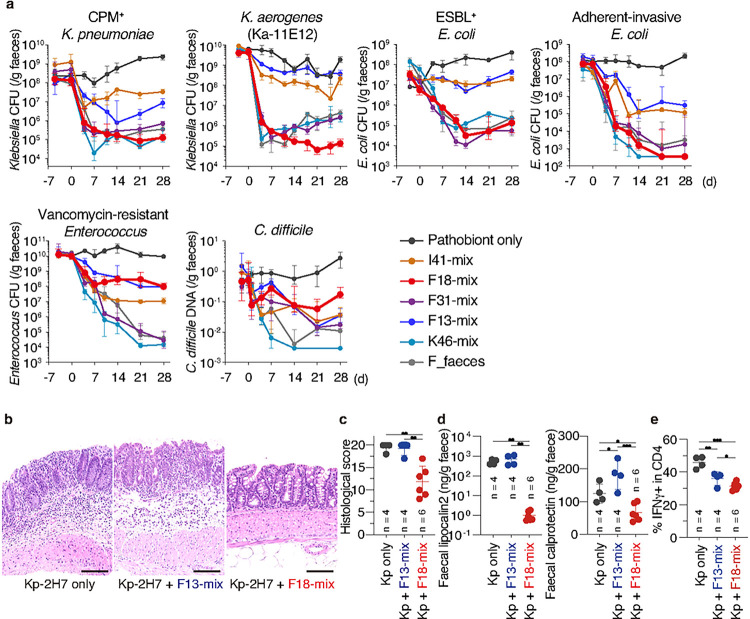
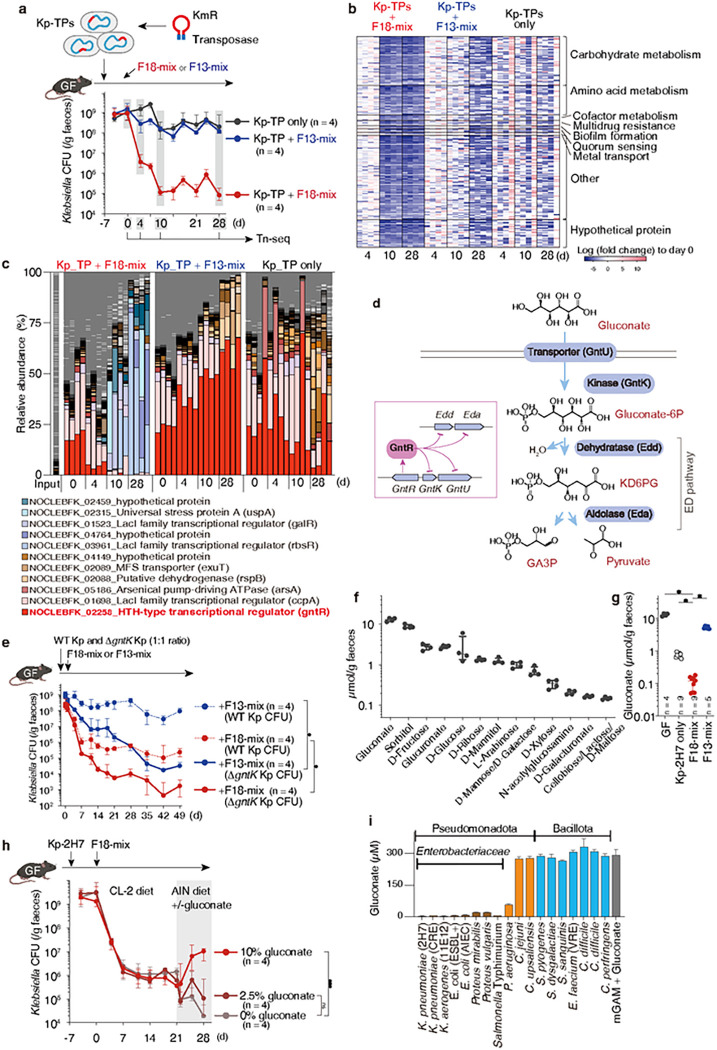
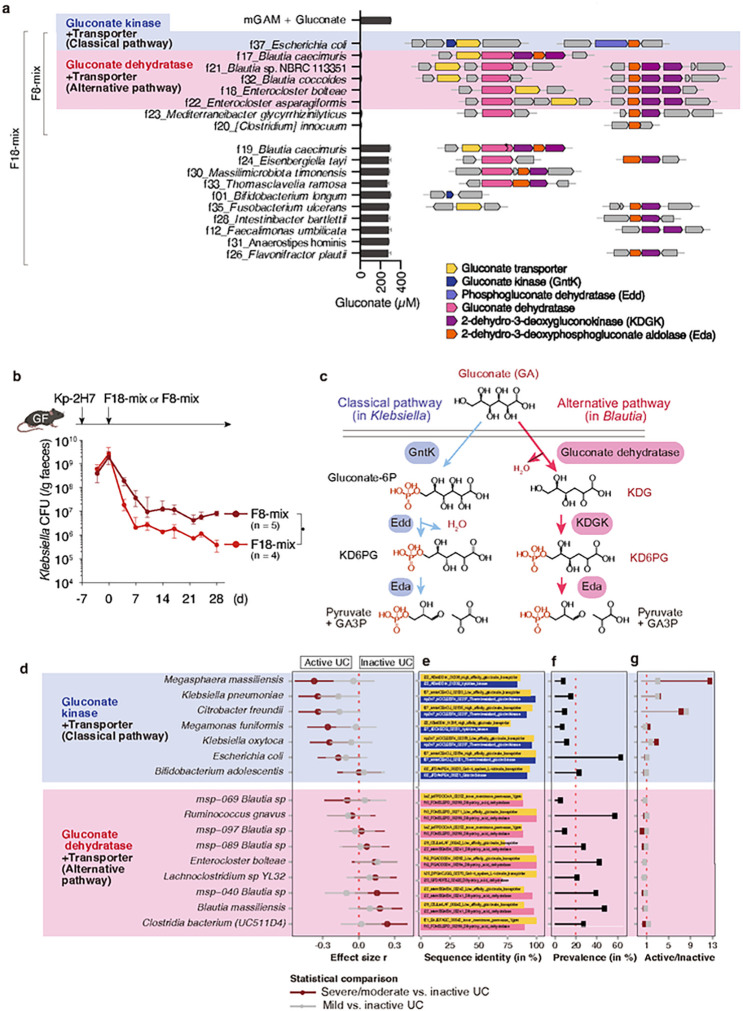
References
-
- Centers for Disease Control and Prevention: Antibiotic Resistance Threats in the United States, 2019. Atlanta, GA, Centers for Disease Control and Prevention. Available at:, <https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-re...> (2019).
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
