

This is a preprint.
Parabrachial Calca neurons drive nociplasticity
- PMID: 37961621
- PMCID: PMC10634894
- DOI: 10.1101/2023.10.26.564223
Parabrachial Calca neurons drive nociplasticity
Update in
-
Parabrachial Calca neurons drive nociplasticity.Cell Rep. 2024 Apr 23;43(4):114057. doi: 10.1016/j.celrep.2024.114057. Epub 2024 Apr 6. Cell Rep. 2024. PMID: 38583149 Free PMC article.
Abstract
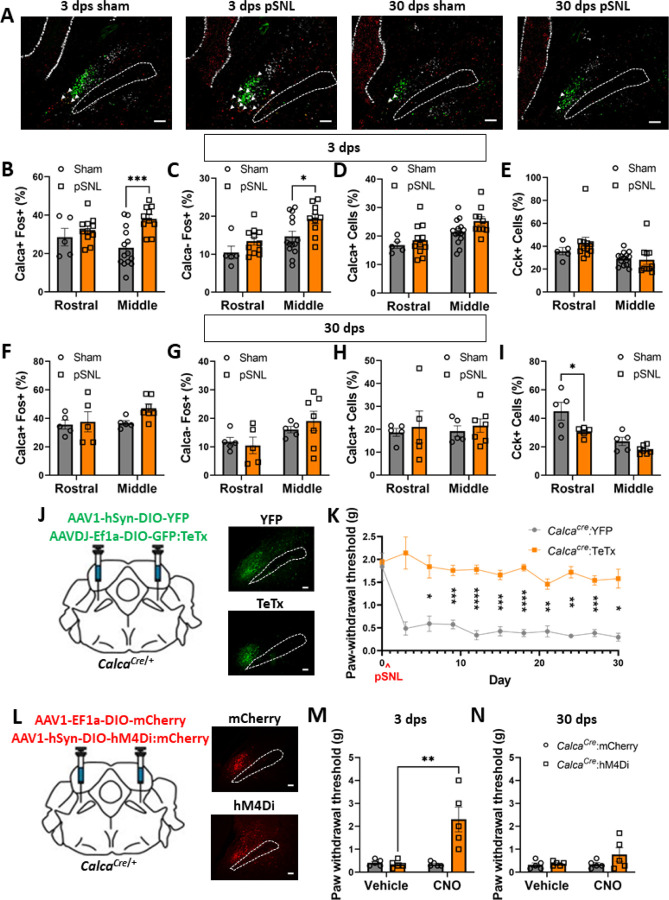
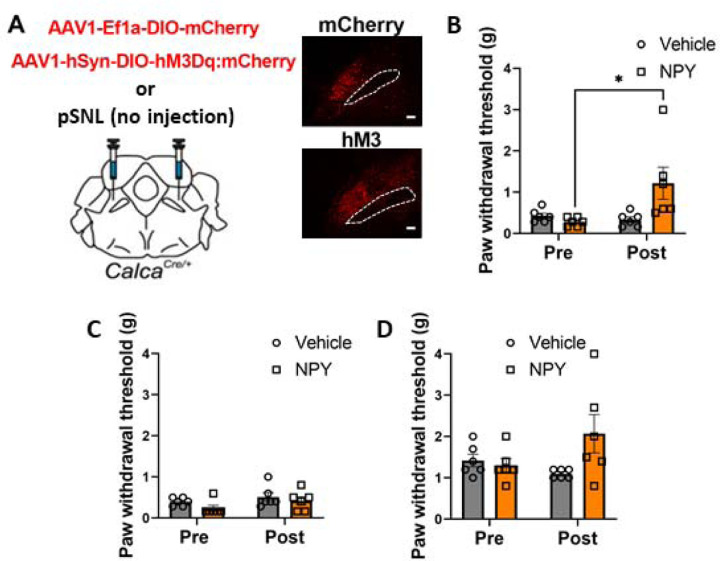
Pain that persists beyond the time required for tissue healing and pain that arises in the absence of tissue injury are poorly understood phenomena mediated by plasticity within the central nervous system. The parabrachial nucleus (PBN) is a hub that relays aversive sensory information and appears to play a role in nociplasticity. Here, by preventing PBN Calca neurons from releasing neurotransmitter or directly stimulating them we demonstrate that activation of Calca neurons is both necessary for the manifestation of chronic pain after nerve ligation and is sufficient to drive nociplasticity in wild-type mice. Aversive stimuli such as exposure to nitroglycerin, cisplatin, or LiCl can drive nociplasticity in a Calca-neuron-dependent manner. Calcium fluorescence imaging reveals that nitroglycerin activates PBN Calca neurons and potentiates their responses to mechanical stimulation. The activity and excitability of Calca neurons increased for several days after aversive events, but prolonged nociplasticity likely occurs in downstream circuitry.
Conflict of interest statement
DECLARARTION OF INTERSTS Authors declare no conflicting interests.
Figures





References
-
- Allen H.N., Chaudhry S., Hong V.M., Lewter L.A., Sinha G.P., Carrasquillo Y., Taylor B.K., Kolber B.J. (2023). A parabrachial-to-amygdala circuit that determines hemispheric lateralization of somatosensory processing. Biol. Psychiatry. 93, 370–381. 10.1016/j.biopsych.2022.09.010. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials