

Formation of an invasion-permissive matrix requires TGFβ/SNAIL1-regulated alternative splicing of fibronectin
- PMID: 37964360
- PMCID: PMC10647173
- DOI: 10.1186/s13058-023-01736-y
Formation of an invasion-permissive matrix requires TGFβ/SNAIL1-regulated alternative splicing of fibronectin
Abstract
Background: As in most solid cancers, the emergence of cells with oncogenic mutations in the mammary epithelium alters the tissue homeostasis. Some soluble factors, such as TGFβ, potently modify the behavior of healthy stromal cells. A subpopulation of cancer-associated fibroblasts expressing a TGFβ target, the SNAIL1 transcription factor, display myofibroblastic abilities that rearrange the stromal architecture. Breast tumors with the presence of SNAIL1 in the stromal compartment, and with aligned extracellular fiber, are associated with poor survival prognoses.
Methods: We used deep RNA sequencing and biochemical techniques to study alternative splicing and human tumor databases to test for associations (correlation t-test) between SNAIL1 and fibronectin isoforms. Three-dimensional extracellular matrices generated from fibroblasts were used to study the mechanical properties and actions of the extracellular matrices on tumor cell and fibroblast behaviors. A metastatic mouse model of breast cancer was used to test the action of fibronectin isoforms on lung metastasis.
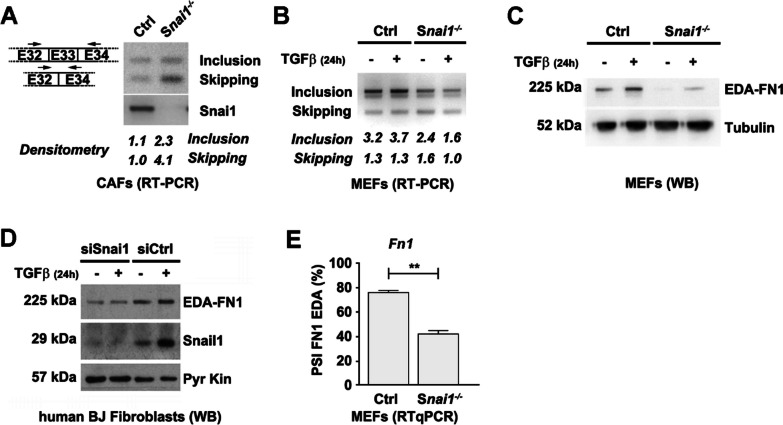
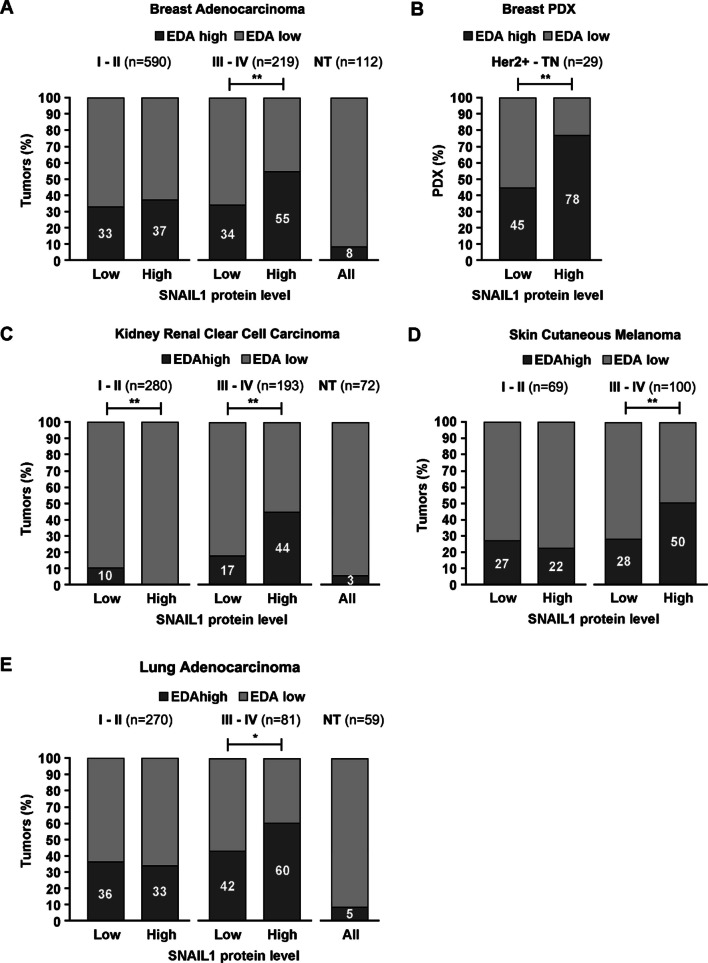
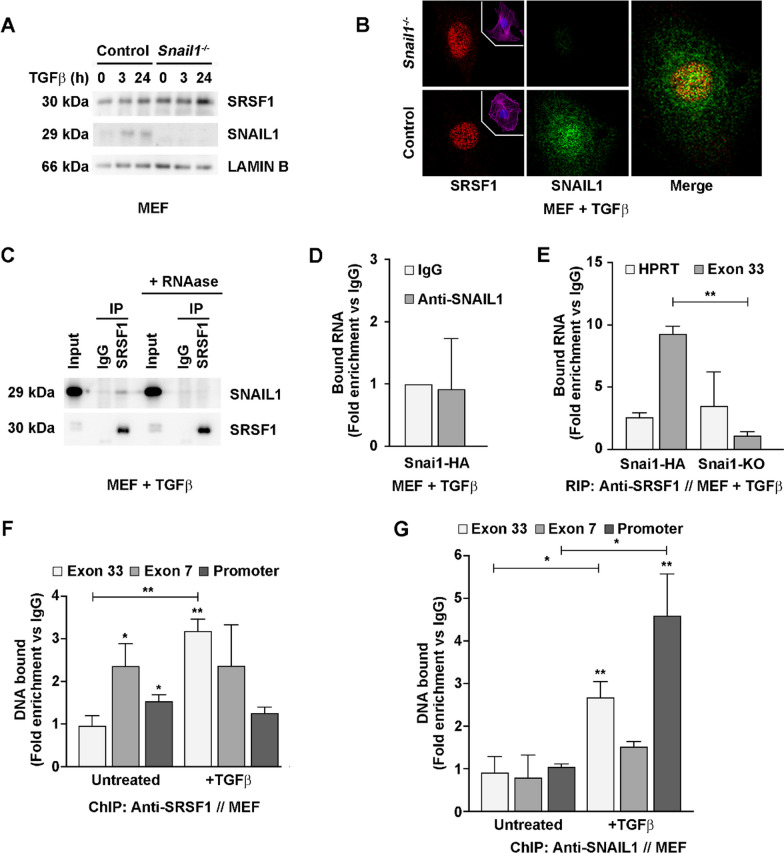
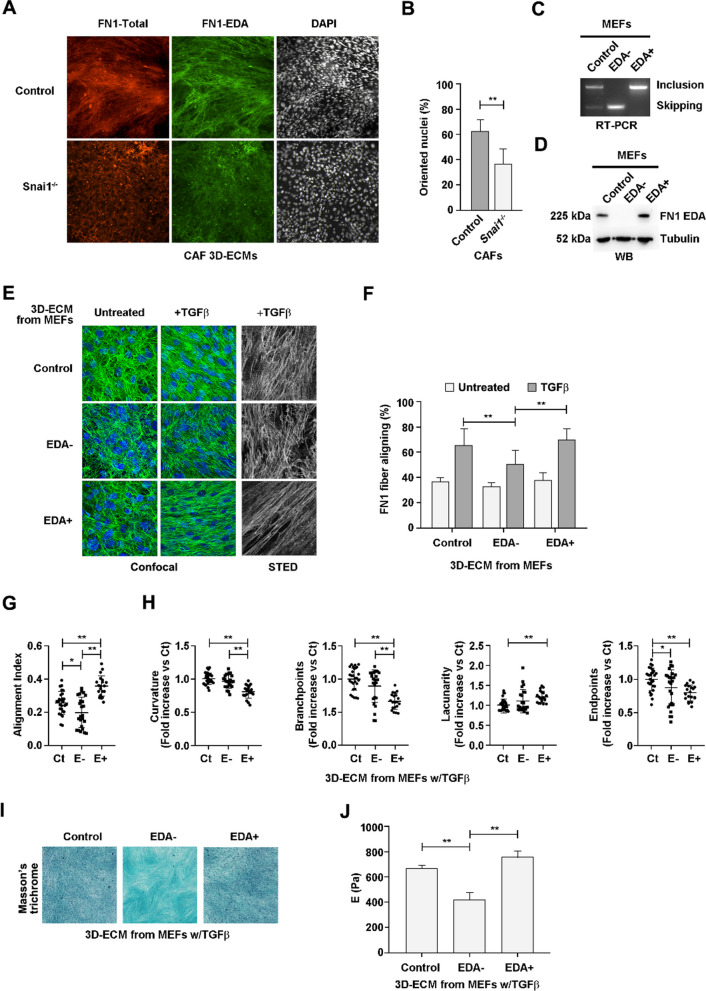
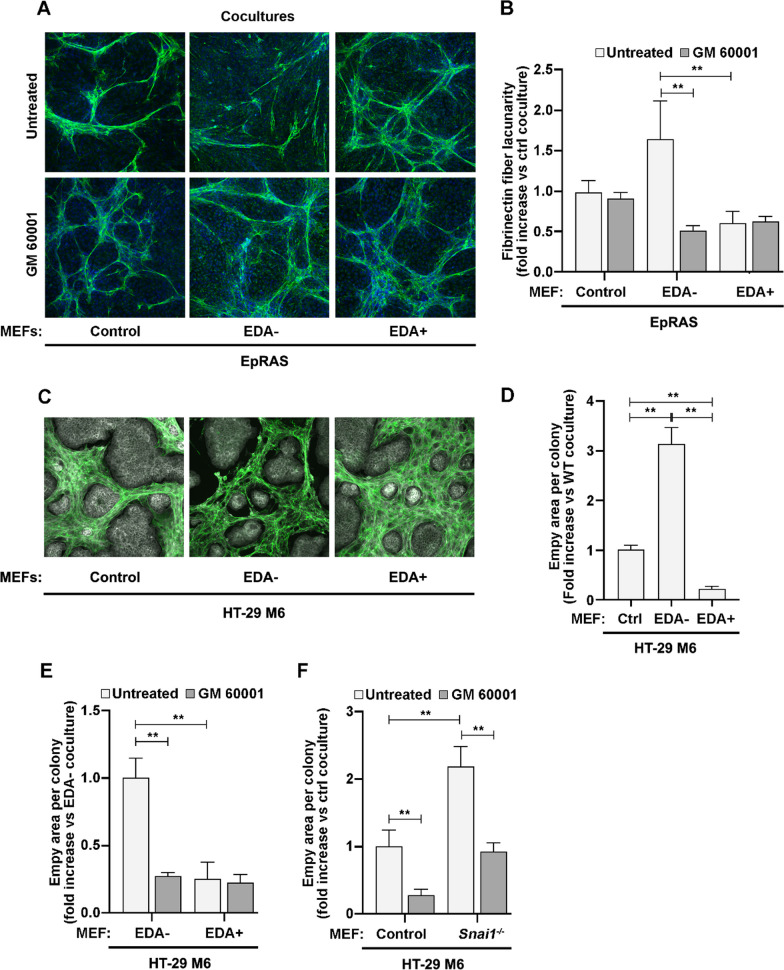
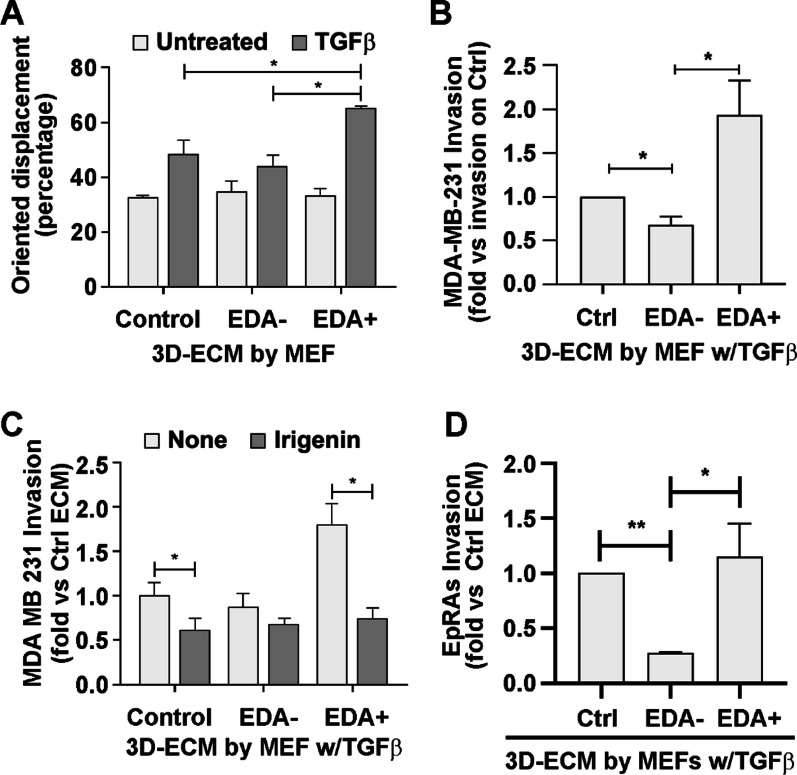
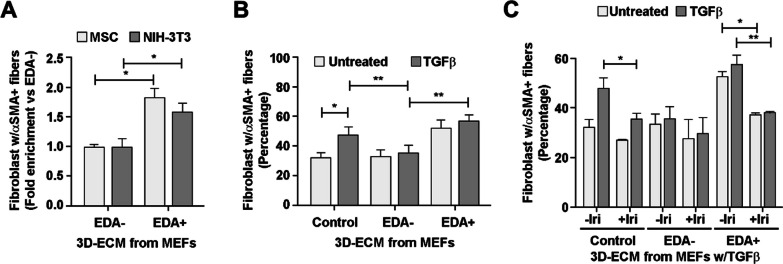
Results: In silico studies showed that SNAIL1 correlates with the expression of the extra domain A (EDA)-containing (EDA+) fibronectin in advanced human breast cancer and other types of epithelial cancers. In TGFβ-activated fibroblasts, alternative splicing of fibronectin as well as of 500 other genes was modified by eliminating SNAIL1. Biochemical analyses demonstrated that SNAIL1 favors the inclusion of the EDA exon by modulating the activity of the SRSF1 splicing factor. Similar to Snai1 knockout fibroblasts, EDA- fibronectin fibroblasts produce an extracellular matrix that does not sustain TGFβ-induced fiber organization, rigidity, fibroblast activation, or tumor cell invasion. The presence of EDA+ fibronectin changes the action of metalloproteinases on fibronectin fibers. Critically, in an mouse orthotopic breast cancer model, the absence of the fibronectin EDA domain completely prevents lung metastasis.
Conclusions: Our results support the requirement of EDA+ fibronectin in the generation of a metastasis permissive stromal architecture in breast cancers and its molecular control by SNAIL1. From a pharmacological point of view, specifically blocking EDA+ fibronectin deposition could be included in studies to reduce the formation of a pro-metastatic environment.
Keywords: Breast cancer; EDA+ Fibronectin; Extracellular matrix; Matrix architecture; Matrix rigidity; Metastasis; Myofibroblasts; SNAIL1; TGFβ.
© 2023. The Author(s).
Conflict of interest statement
The authors have no conflicts of interest to disclosure.
Figures

Similar articles
-
Targeting alternative splicing of fibronectin in human renal proximal tubule epithelial cells with antisense oligonucleotides to reduce EDA+ fibronectin production and block an autocrine loop that drives renal fibrosis.Exp Cell Res. 2024 Sep 1;442(1):114186. doi: 10.1016/j.yexcr.2024.114186. Epub 2024 Aug 2. Exp Cell Res. 2024. PMID: 39098465
-
Alternatively spliced isoforms of fibronectin in immune-mediated glomerulosclerosis: the role of TGFbeta and IL-4.J Pathol. 2004 Nov;204(3):248-57. doi: 10.1002/path.1653. J Pathol. 2004. PMID: 15372454
-
Control of fibroblast fibronectin expression and alternative splicing via the PI3K/Akt/mTOR pathway.Exp Cell Res. 2010 Oct 1;316(16):2644-53. doi: 10.1016/j.yexcr.2010.06.028. Epub 2010 Jul 13. Exp Cell Res. 2010. PMID: 20615404 Free PMC article.
-
Fibronectin-targeted drug delivery in cancer.Adv Drug Deliv Rev. 2016 Feb 1;97:101-10. doi: 10.1016/j.addr.2015.11.014. Epub 2015 Nov 27. Adv Drug Deliv Rev. 2016. PMID: 26639577 Review.
-
New insights into form and function of fibronectin splice variants.J Pathol. 2008 Sep;216(1):1-14. doi: 10.1002/path.2388. J Pathol. 2008. PMID: 18680111 Free PMC article. Review.
Cited by
-
Shaping Oncogenic Microenvironments: Contribution of Fibronectin.Front Cell Dev Biol. 2024 Apr 10;12:1363004. doi: 10.3389/fcell.2024.1363004. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 38660622 Free PMC article. Review.
-
Best evidence linking the extracellular factor TGF-β to cancer-associated alternative splicing programs.BBA Adv. 2024 Dec 18;7:100132. doi: 10.1016/j.bbadva.2024.100132. eCollection 2025. BBA Adv. 2024. PMID: 40585016 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
