

Disruption of CFAP418 interaction with lipids causes widespread abnormal membrane-associated cellular processes in retinal degenerations
- PMID: 37971880
- PMCID: PMC10906455
- DOI: 10.1172/jci.insight.162621
Disruption of CFAP418 interaction with lipids causes widespread abnormal membrane-associated cellular processes in retinal degenerations
Abstract
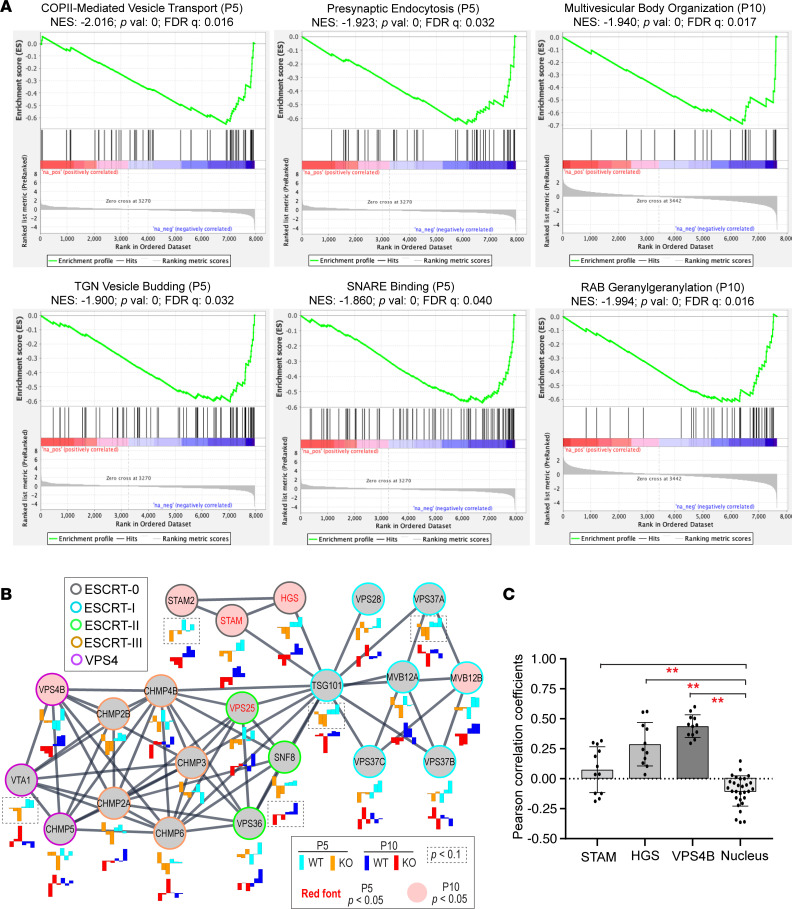
Syndromic ciliopathies and retinal degenerations are large heterogeneous groups of genetic diseases. Pathogenic variants in the CFAP418 gene may cause both disorders, and its protein sequence is evolutionarily conserved. However, the disease mechanism underlying CFAP418 mutations has not been explored. Here, we apply quantitative lipidomic, proteomic, and phosphoproteomic profiling and affinity purification coupled with mass spectrometry to address the molecular function of CFAP418 in the retina. We show that CFAP418 protein binds to the lipid metabolism precursor phosphatidic acid (PA) and mitochondrion-specific lipid cardiolipin but does not form a tight and static complex with proteins. Loss of Cfap418 in mice disturbs membrane lipid homeostasis and membrane-protein associations, which subsequently causes mitochondrial defects and membrane-remodeling abnormalities across multiple vesicular trafficking pathways in photoreceptors, especially the endosomal sorting complexes required for transport (ESCRT) pathway. Ablation of Cfap418 also increases the activity of PA-binding protein kinase Cα in the retina. Overall, our results indicate that membrane lipid imbalance is a pathological mechanism underlying syndromic ciliopathies and retinal degenerations which is associated with other known causative genes of these diseases.
Keywords: Cell Biology; Ophthalmology; Protein traffic; Proteomics; Retinopathy.
Conflict of interest statement
Figures








Similar articles
-
Aberrant protein trafficking in retinal degenerations: The initial phase of retinal remodeling.Exp Eye Res. 2016 Sep;150:71-80. doi: 10.1016/j.exer.2015.11.007. Epub 2015 Nov 26. Exp Eye Res. 2016. PMID: 26632497 Free PMC article. Review.
-
Mapping membrane lipids in the developing and adult mouse retina under physiological and pathological conditions using mass spectrometry.J Biol Chem. 2021 Jan-Jun;296:100303. doi: 10.1016/j.jbc.2021.100303. Epub 2021 Jan 16. J Biol Chem. 2021. PMID: 33465374 Free PMC article.
-
Retinal ciliopathies through the lens of Bardet-Biedl Syndrome: Past, present and future.Prog Retin Eye Res. 2022 Jul;89:101035. doi: 10.1016/j.preteyeres.2021.101035. Epub 2021 Dec 18. Prog Retin Eye Res. 2022. PMID: 34929400 Review.
-
Non-syndromic retinal ciliopathies: translating gene discovery into therapy.Hum Mol Genet. 2012 Oct 15;21(R1):R111-24. doi: 10.1093/hmg/dds298. Epub 2012 Jul 26. Hum Mol Genet. 2012. PMID: 22843501 Review.
-
Ion-current-based proteomic profiling of the retina in a rat model of Smith-Lemli-Opitz syndrome.Mol Cell Proteomics. 2013 Dec;12(12):3583-98. doi: 10.1074/mcp.M113.027847. Epub 2013 Aug 26. Mol Cell Proteomics. 2013. PMID: 23979708 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
