

Post-translational modification by the Pgf glycosylation machinery modulates Streptococcus mutans OMZ175 physiology and virulence
- PMID: 37972006
- PMCID: PMC11096274
- DOI: 10.1111/mmi.15190
Post-translational modification by the Pgf glycosylation machinery modulates Streptococcus mutans OMZ175 physiology and virulence
Abstract
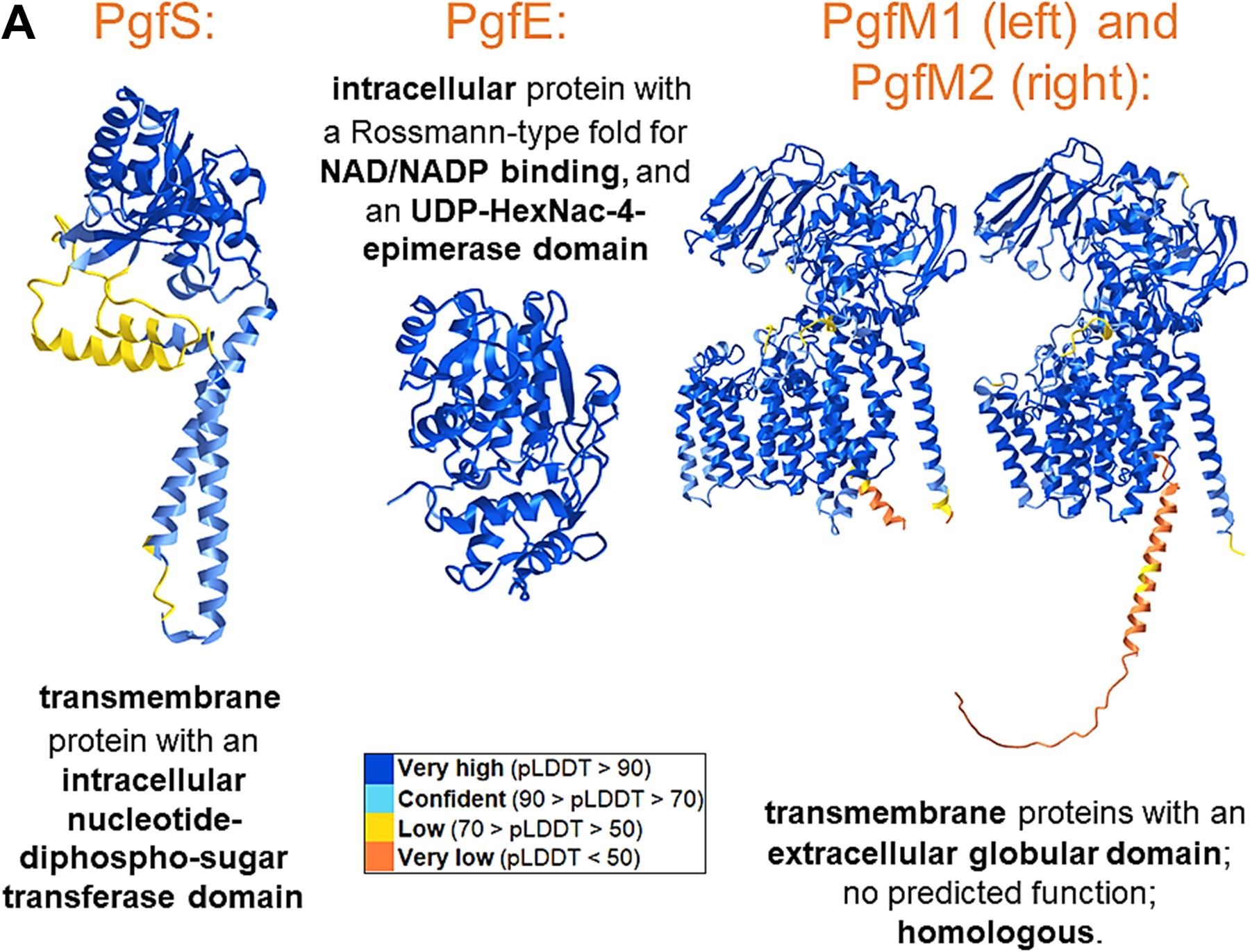
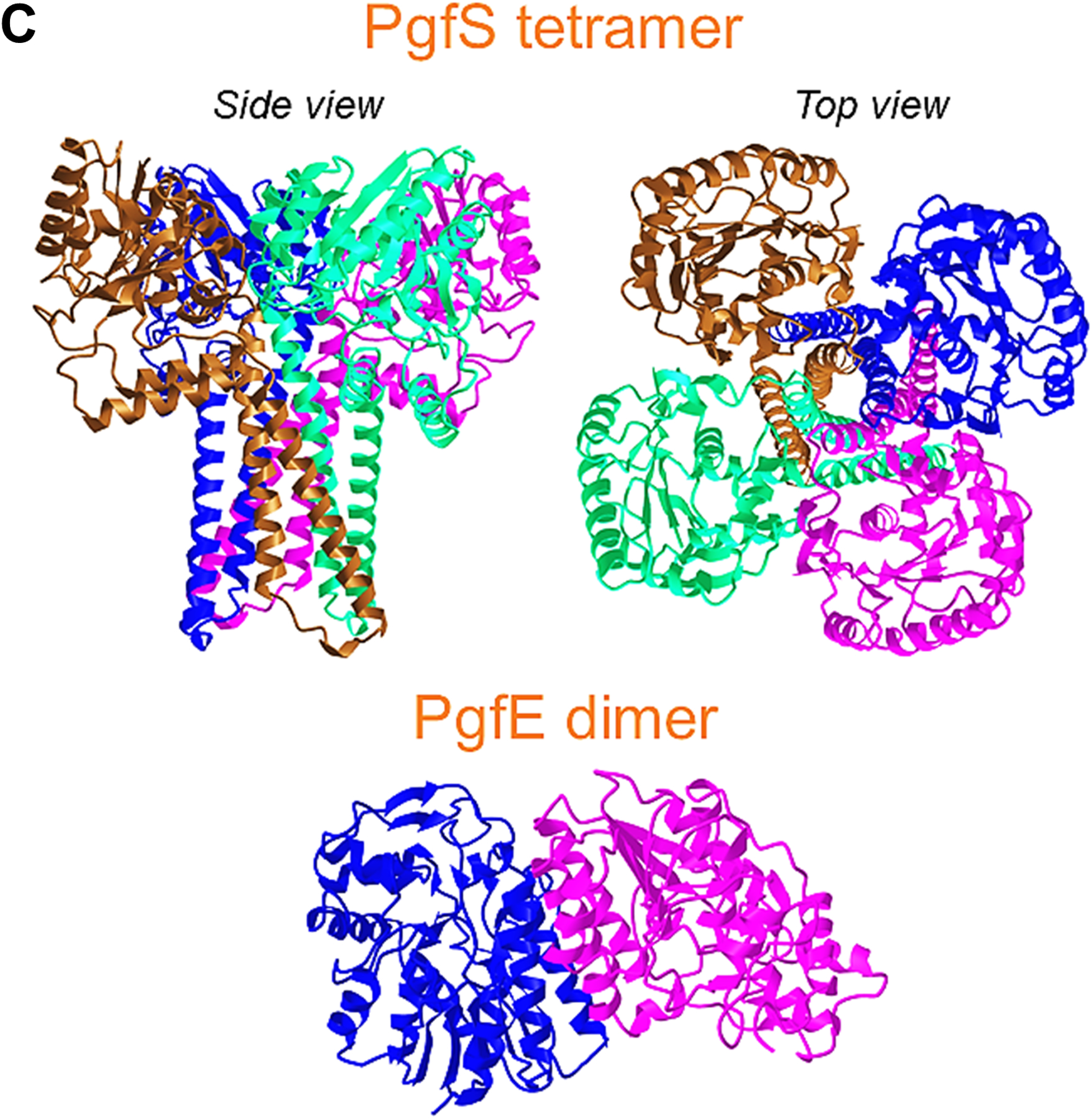
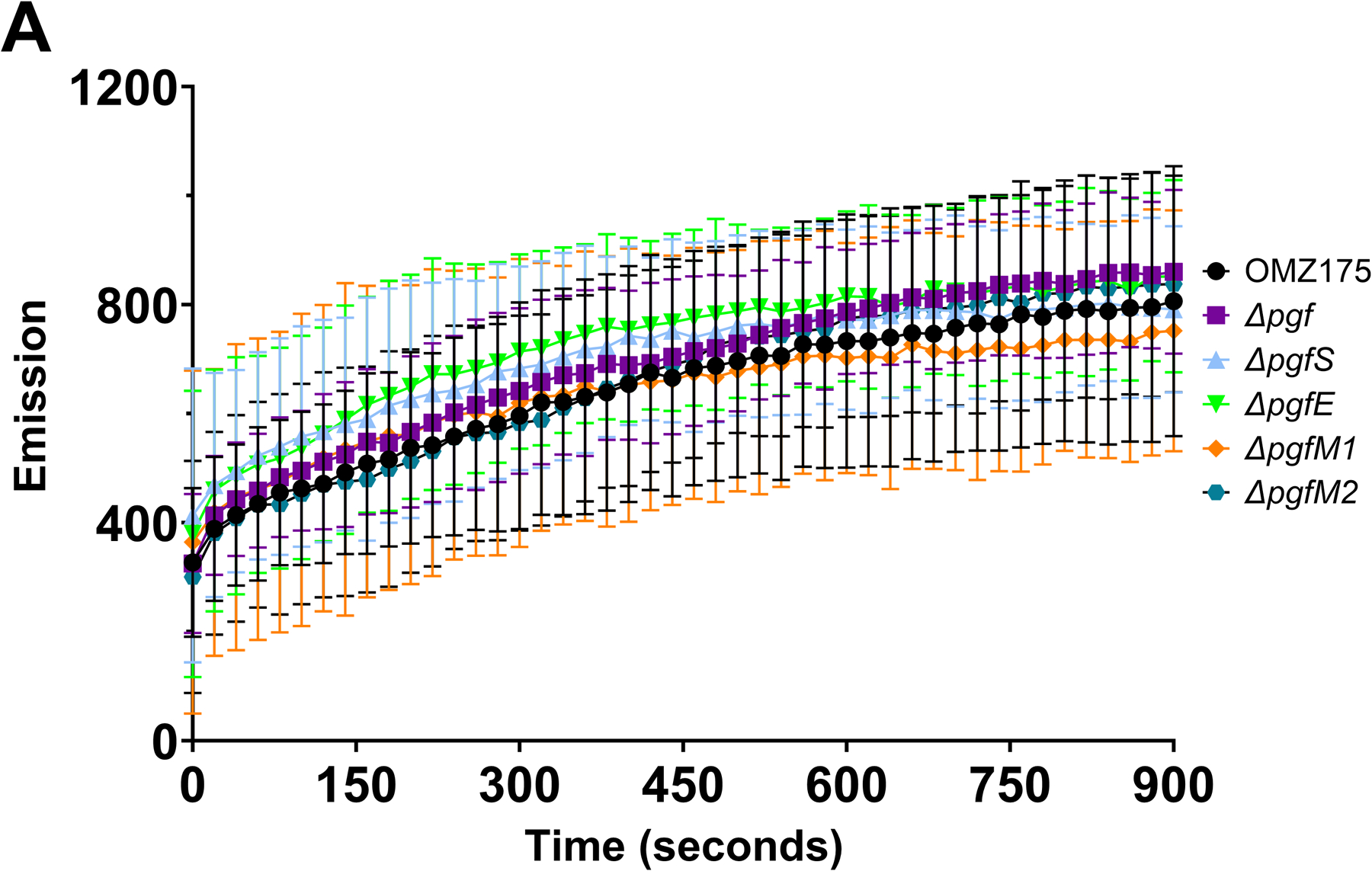
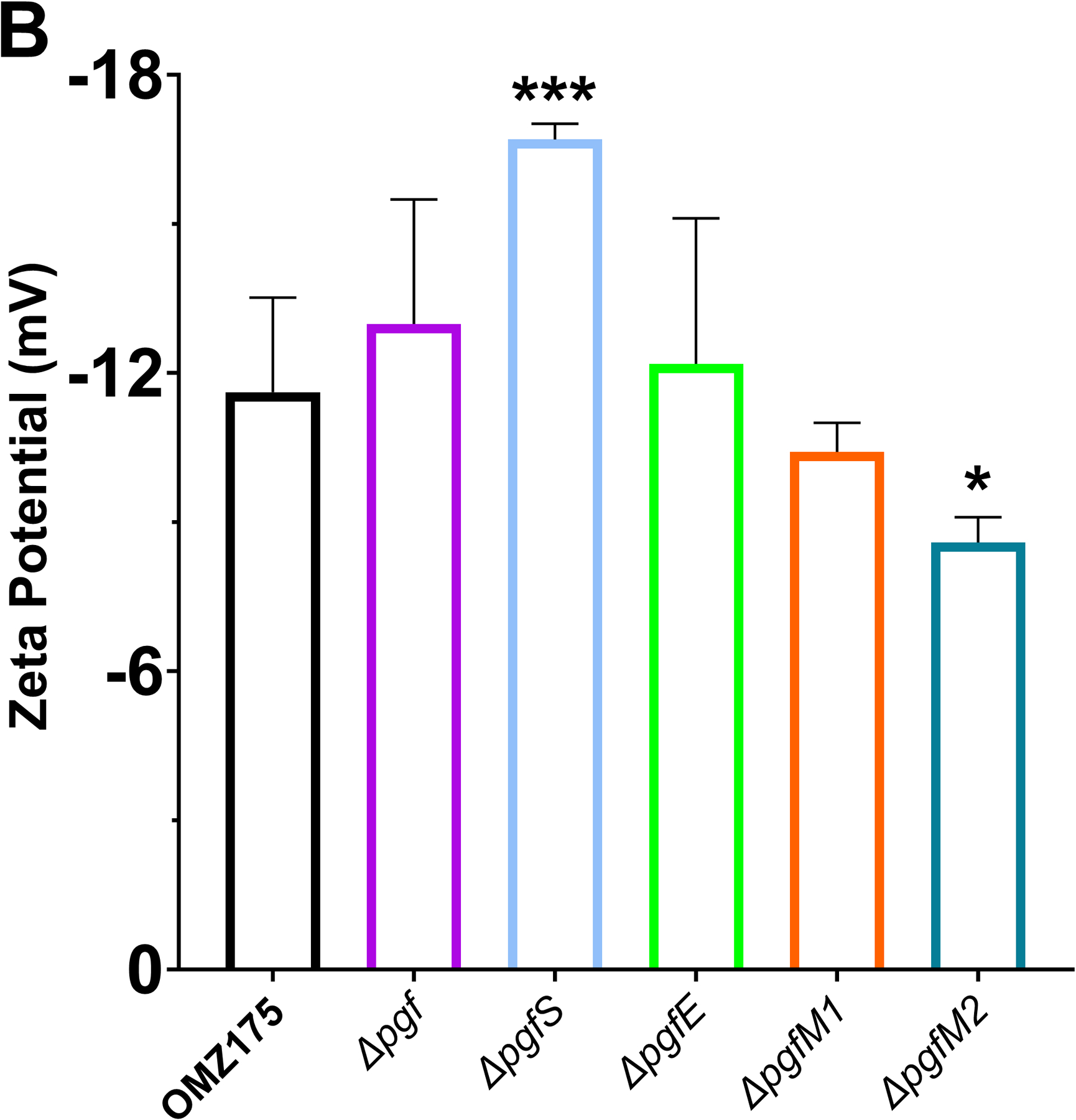
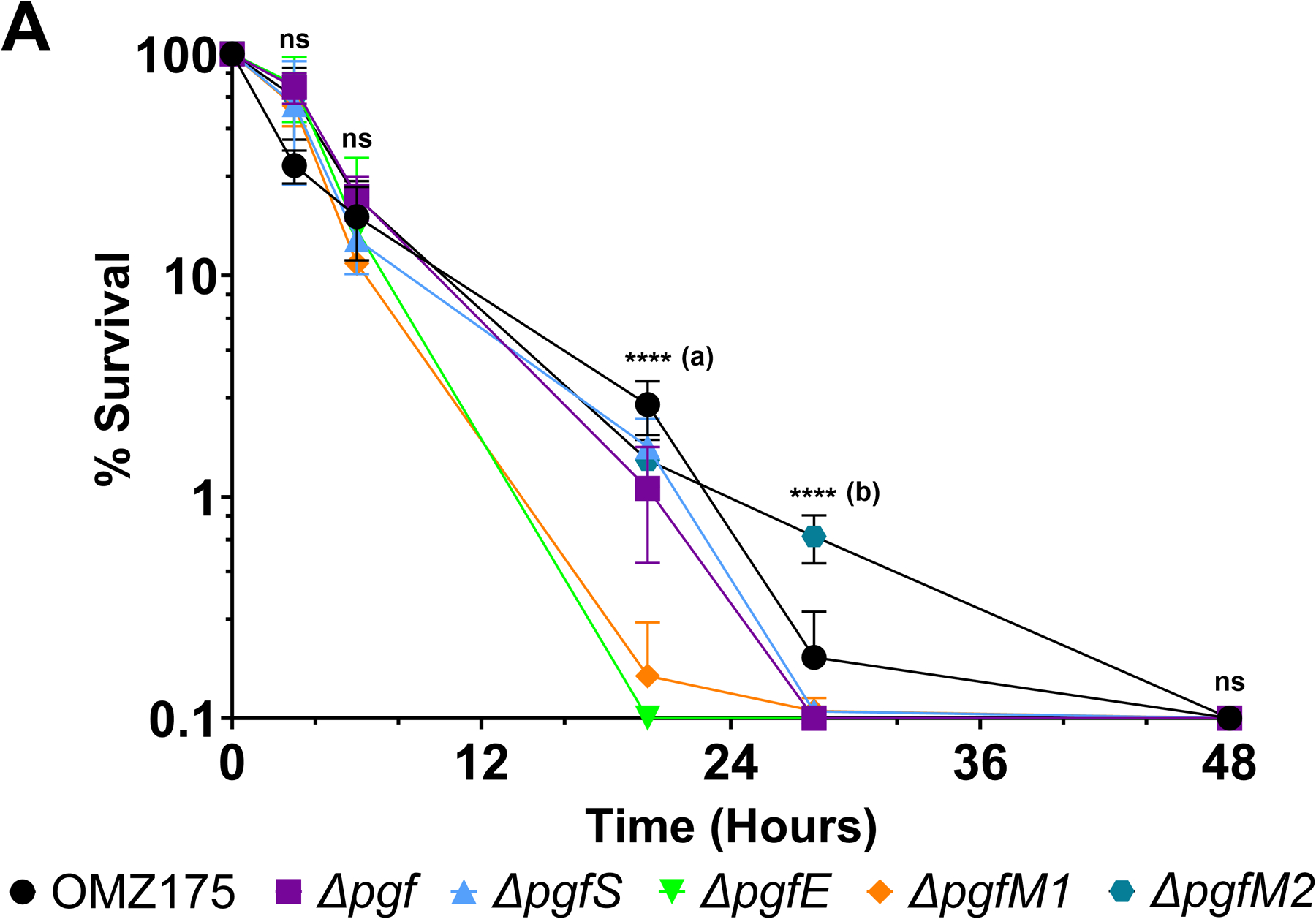
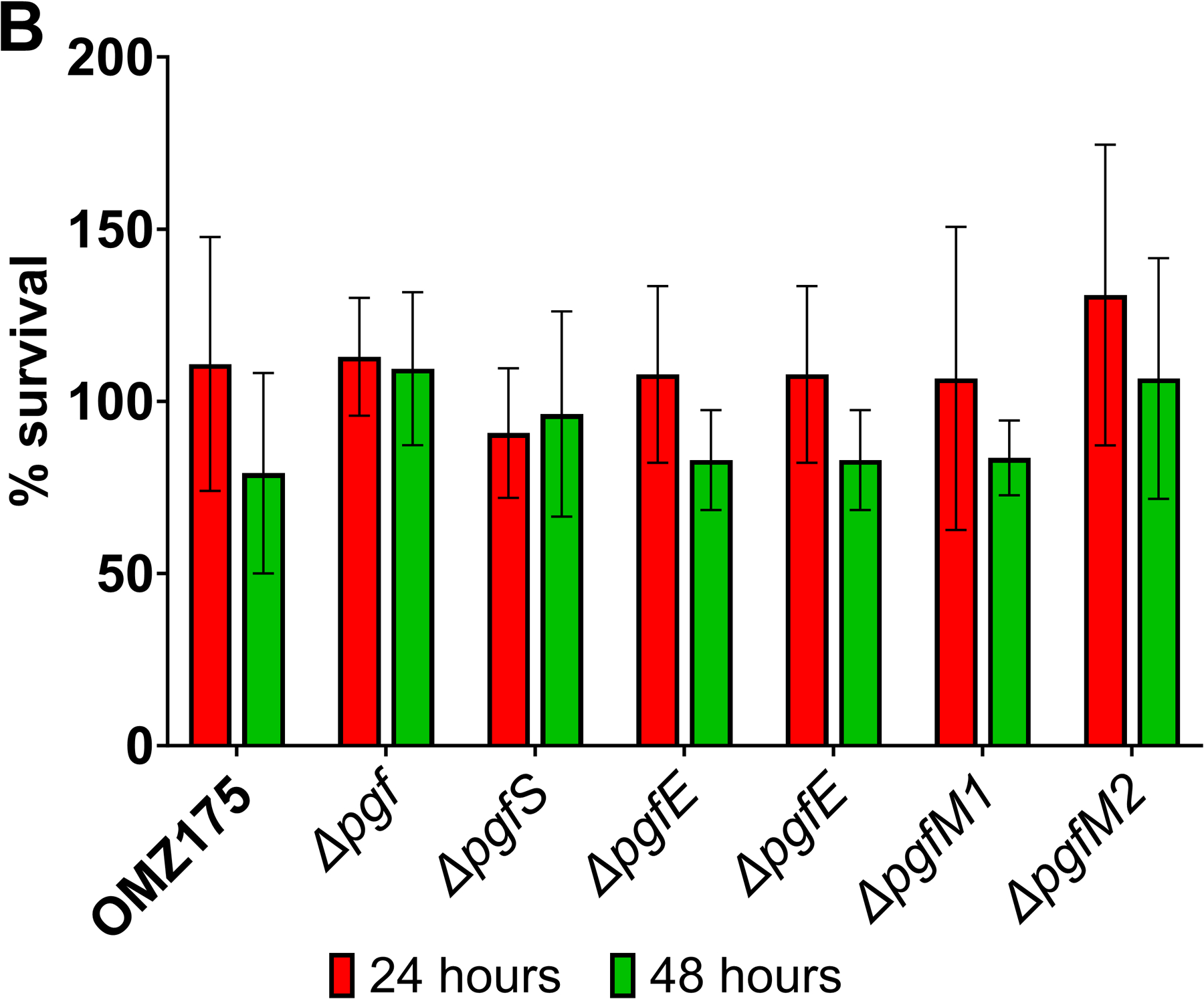
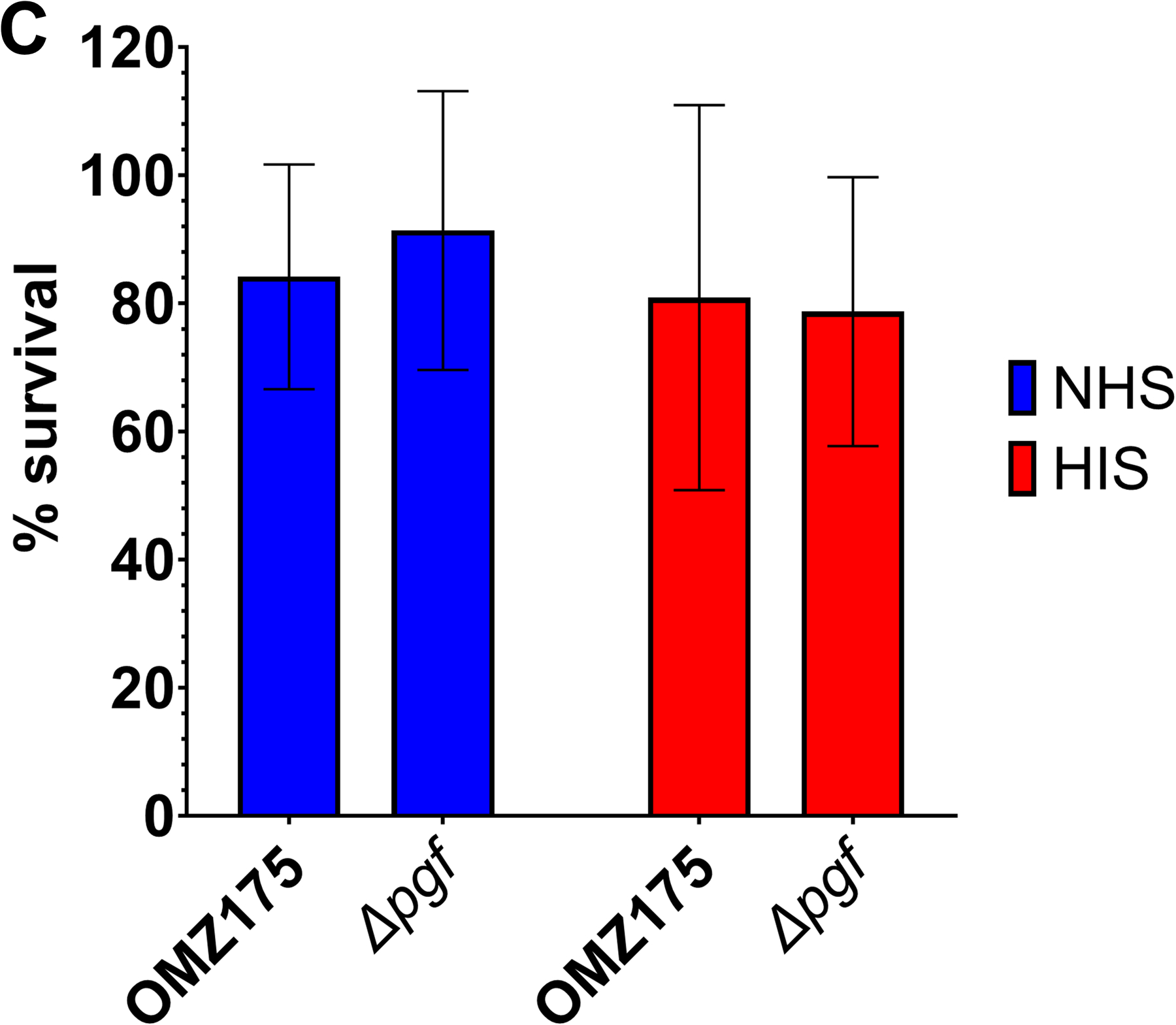
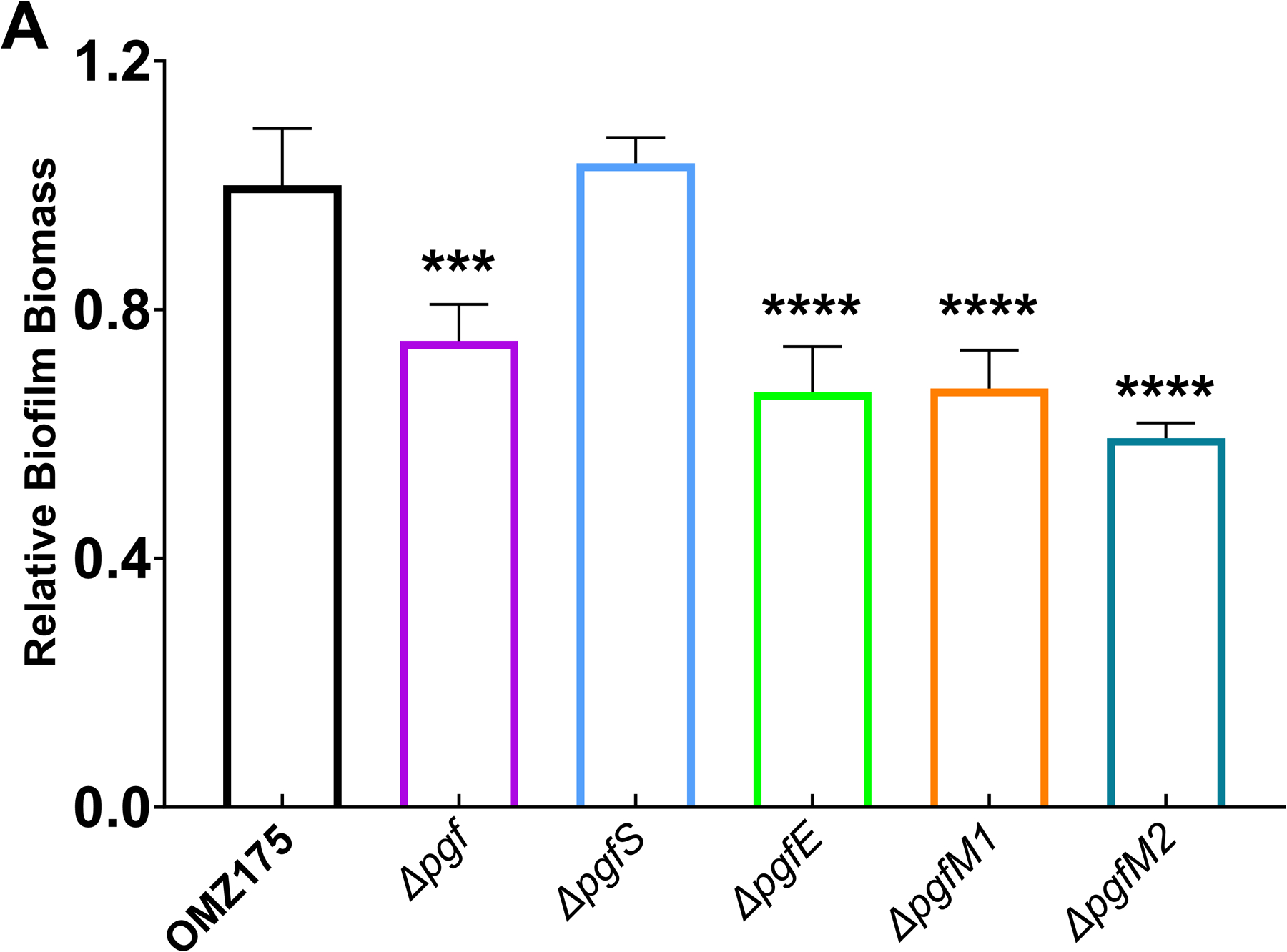
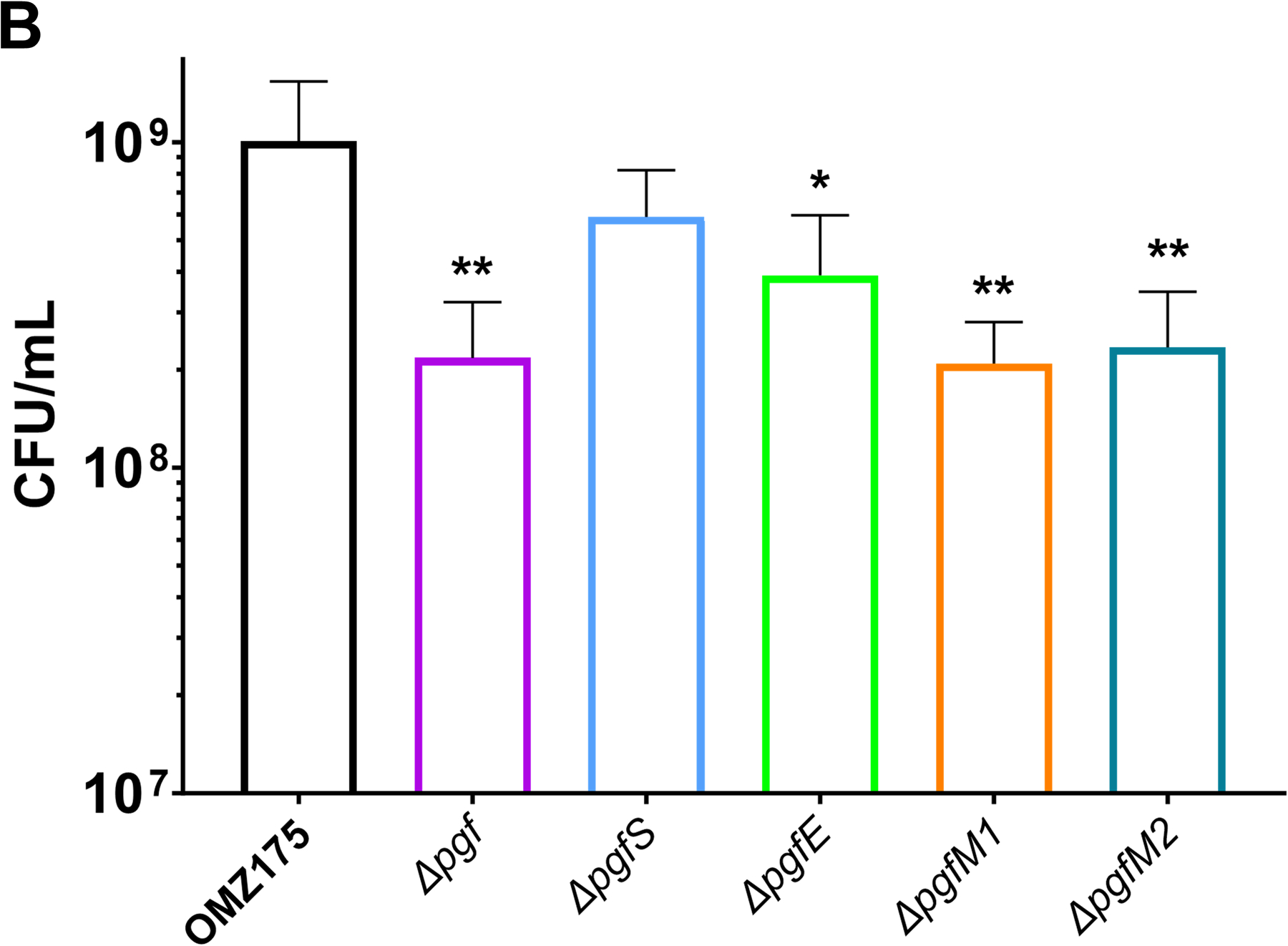
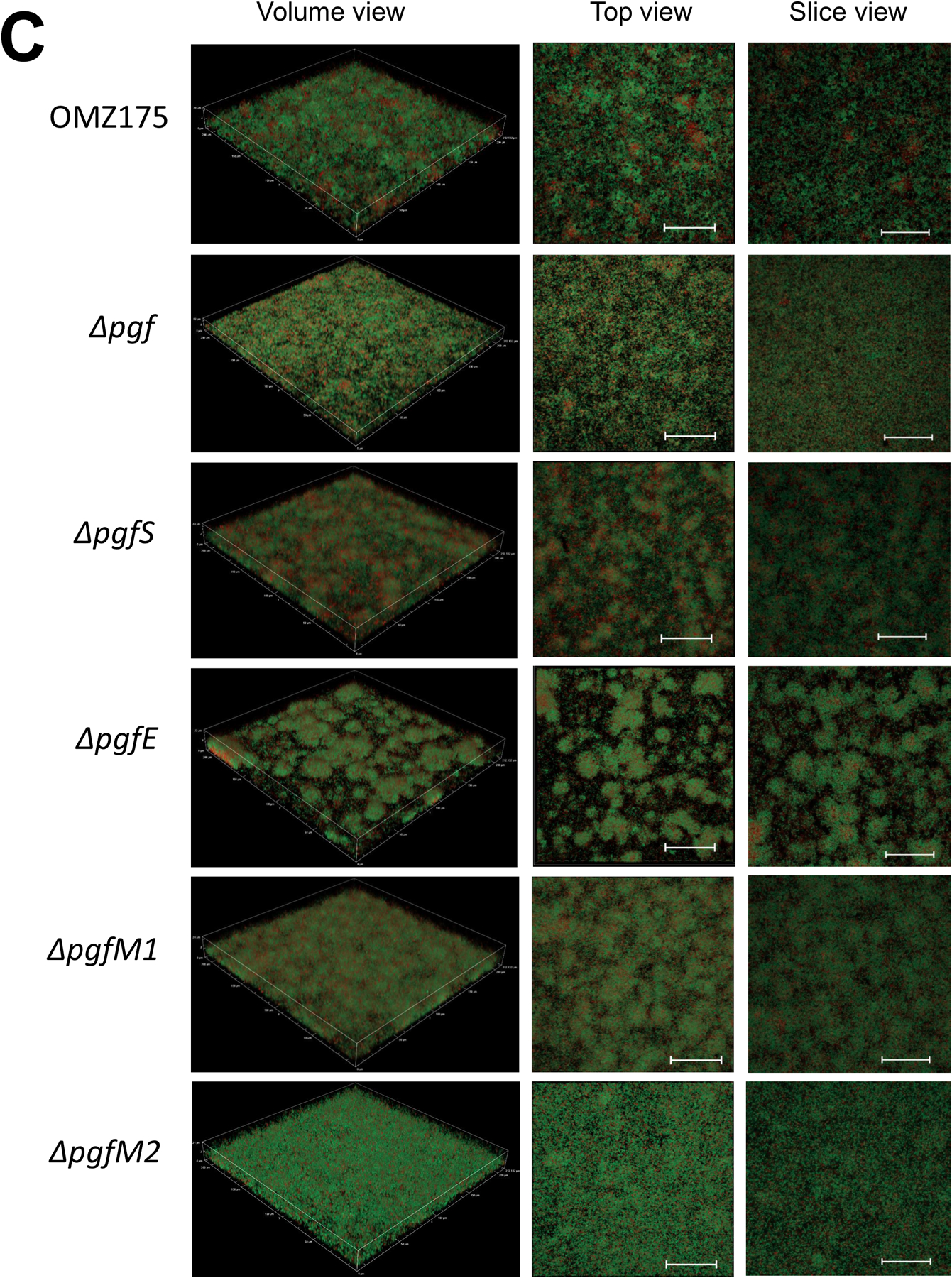
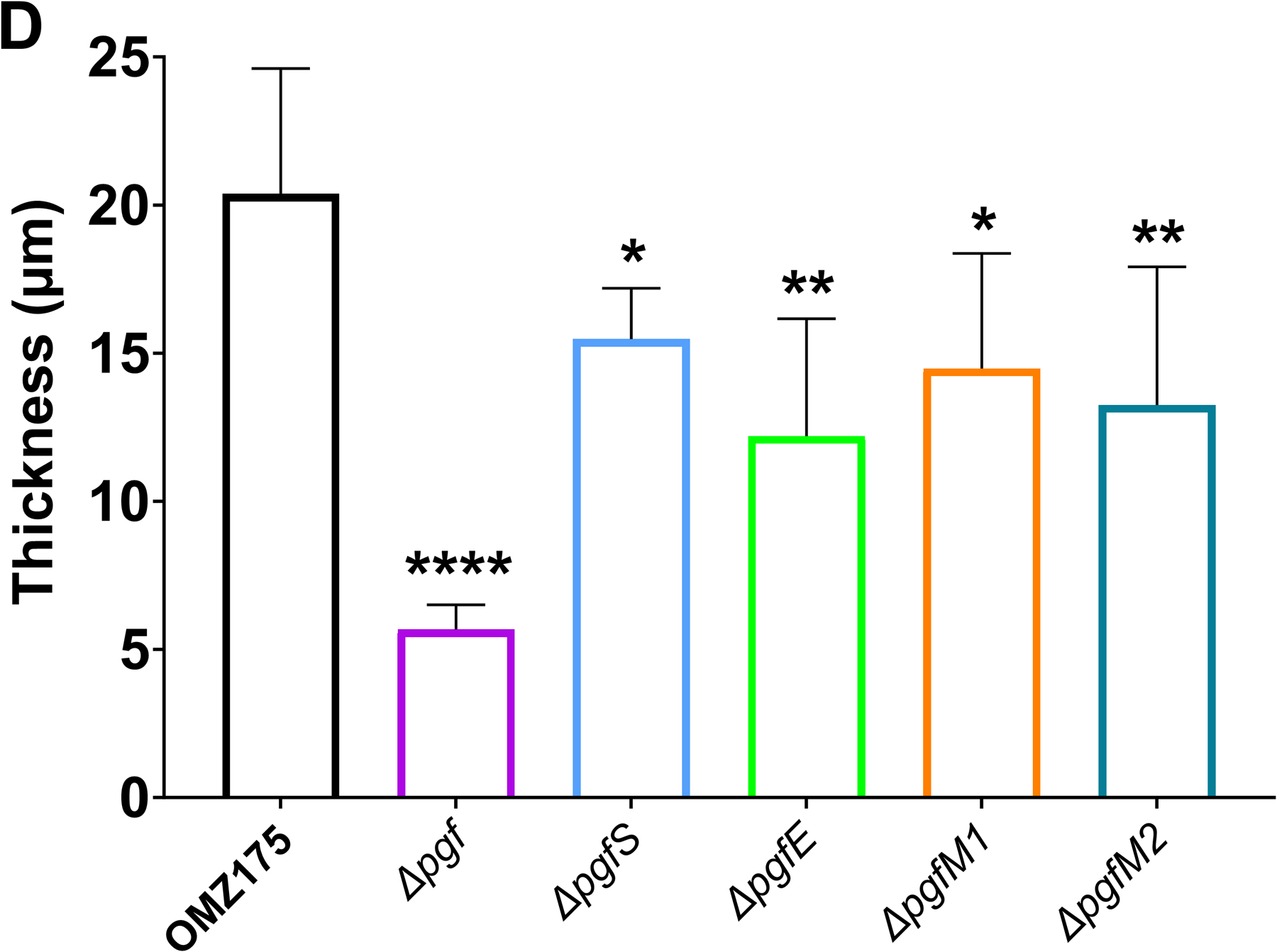
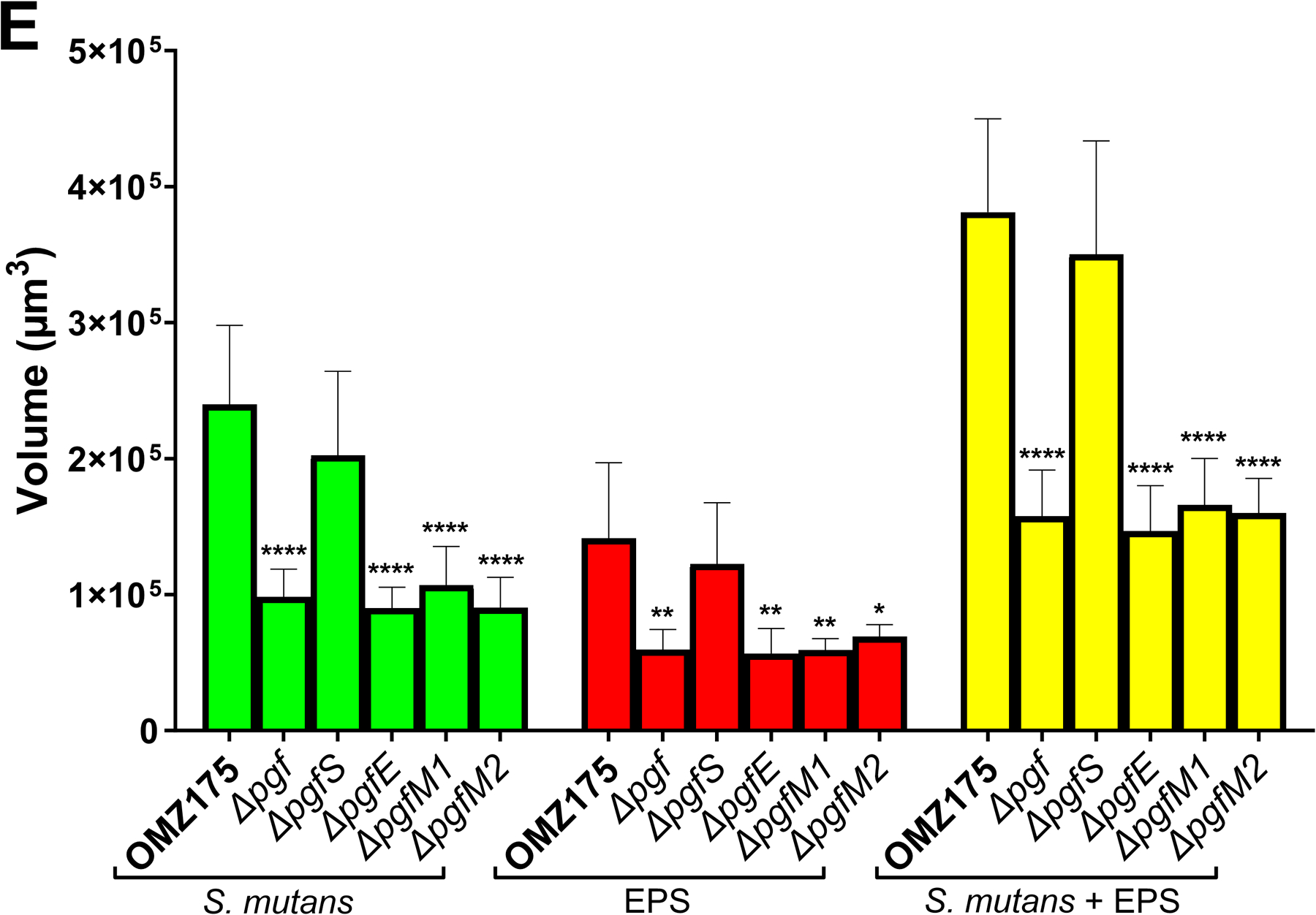
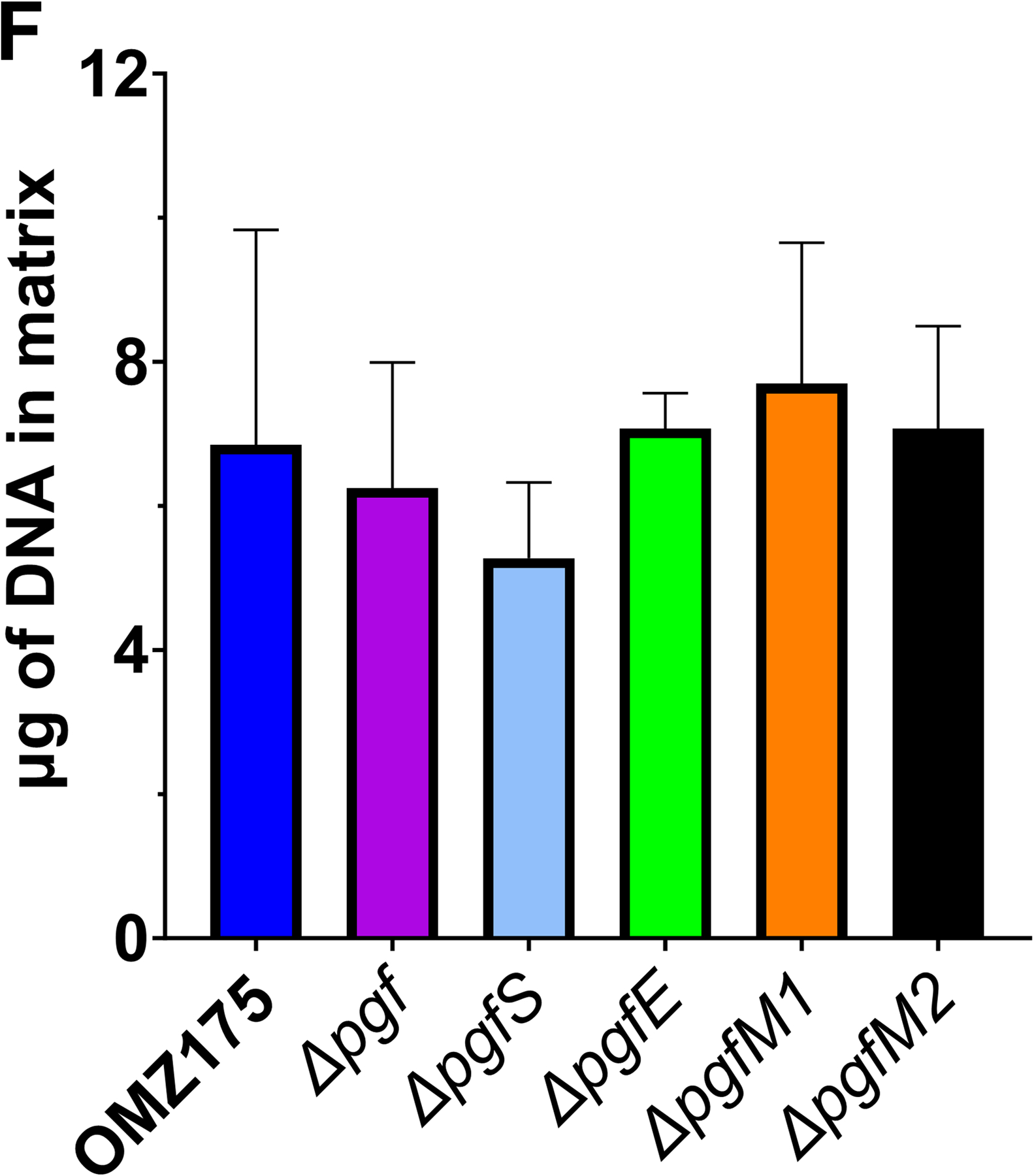
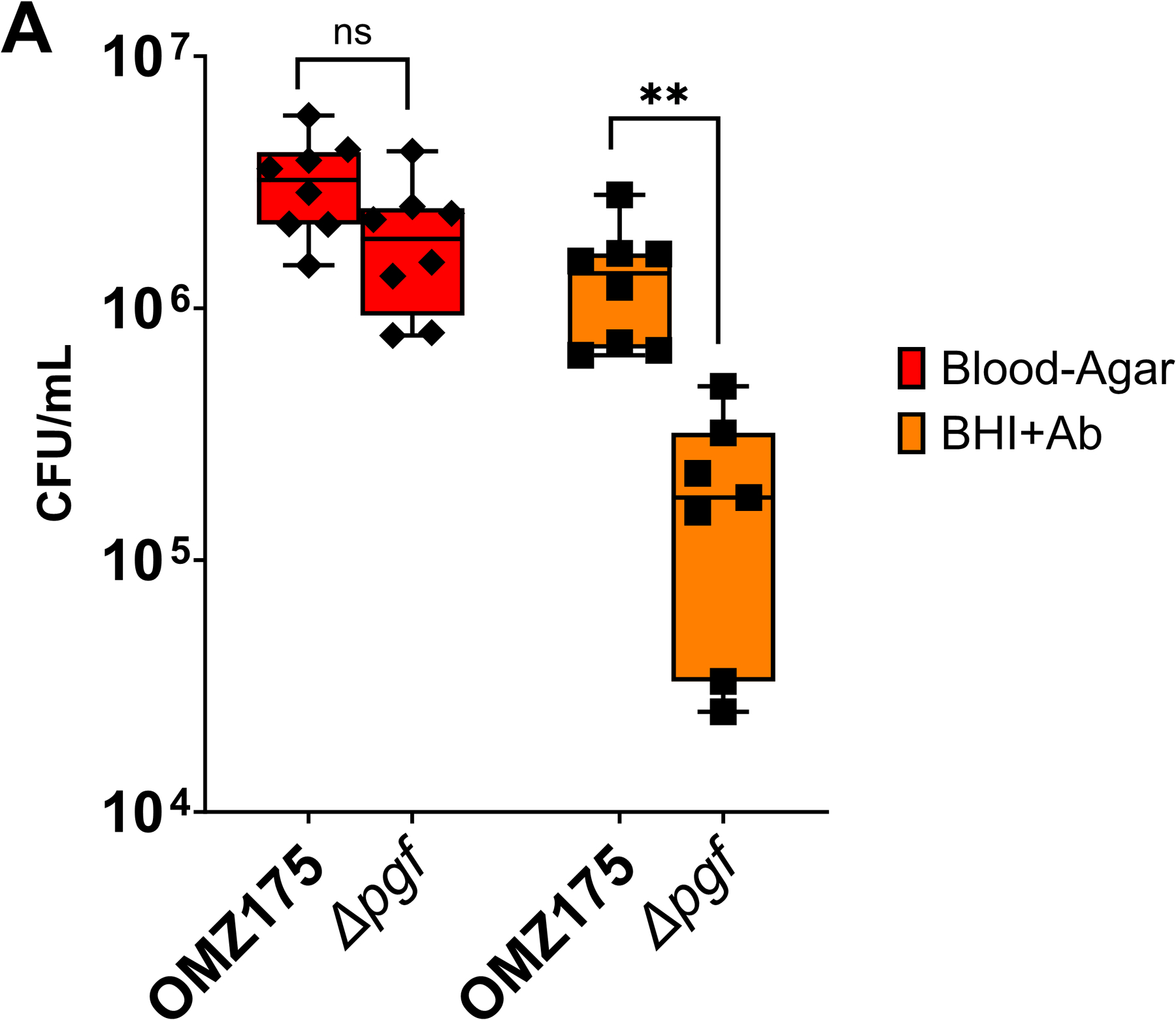
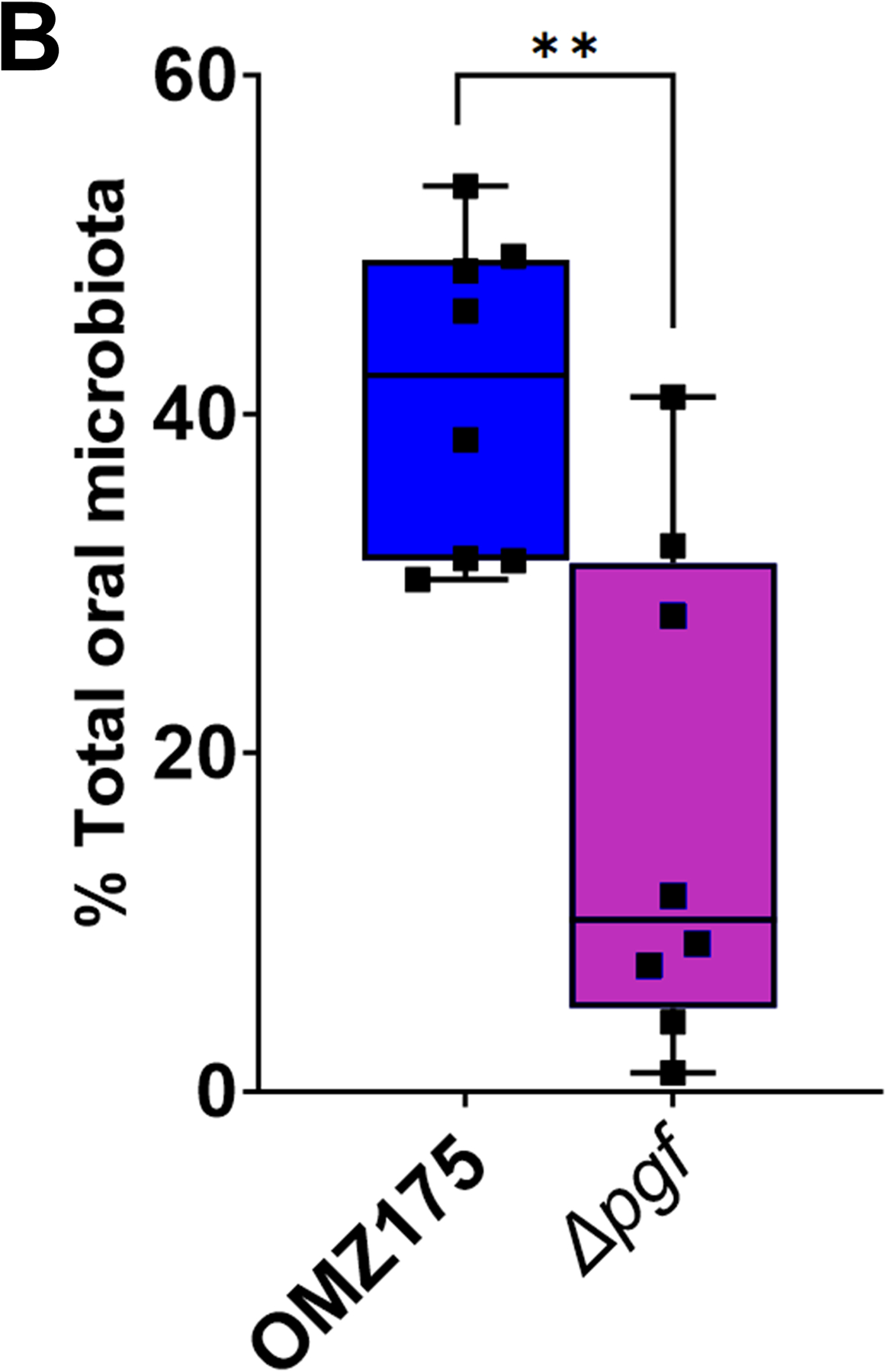
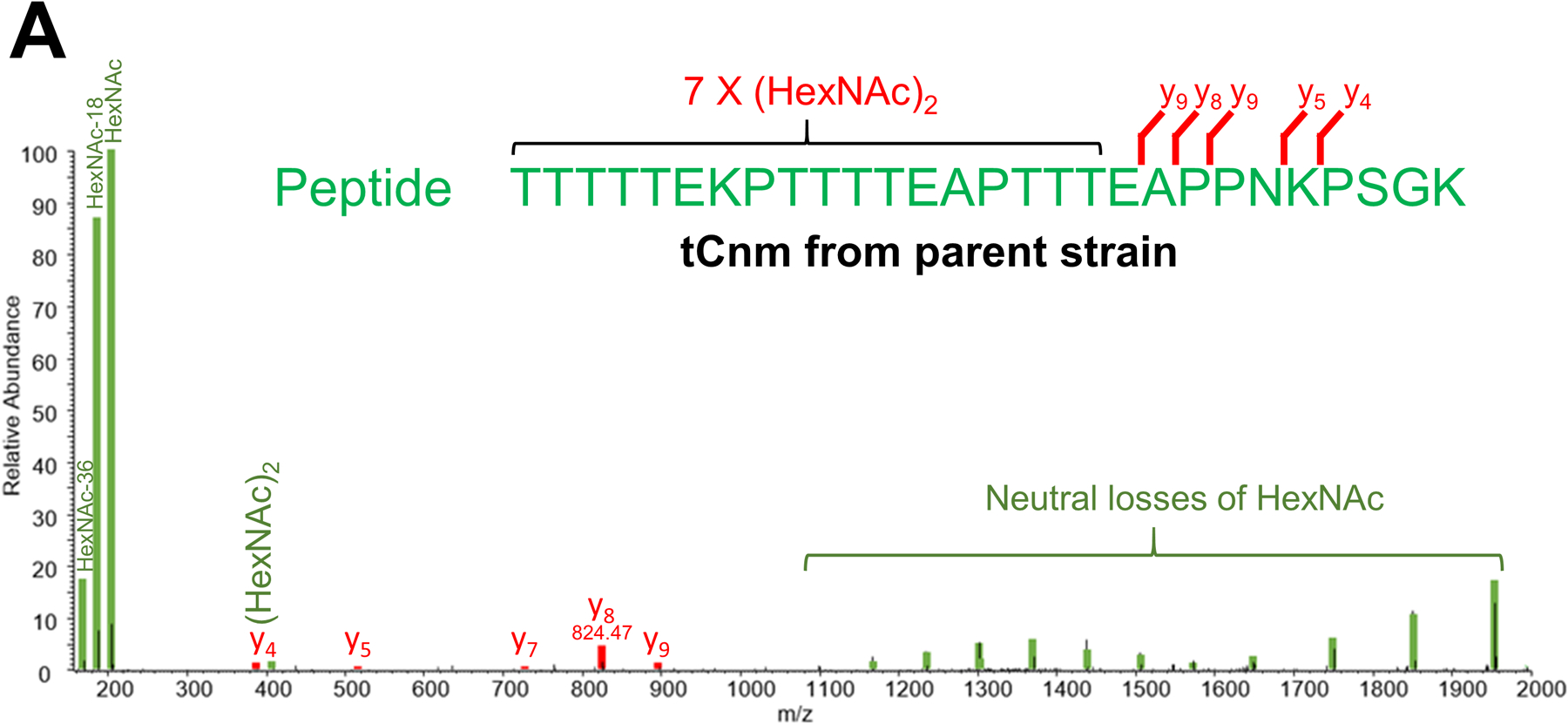
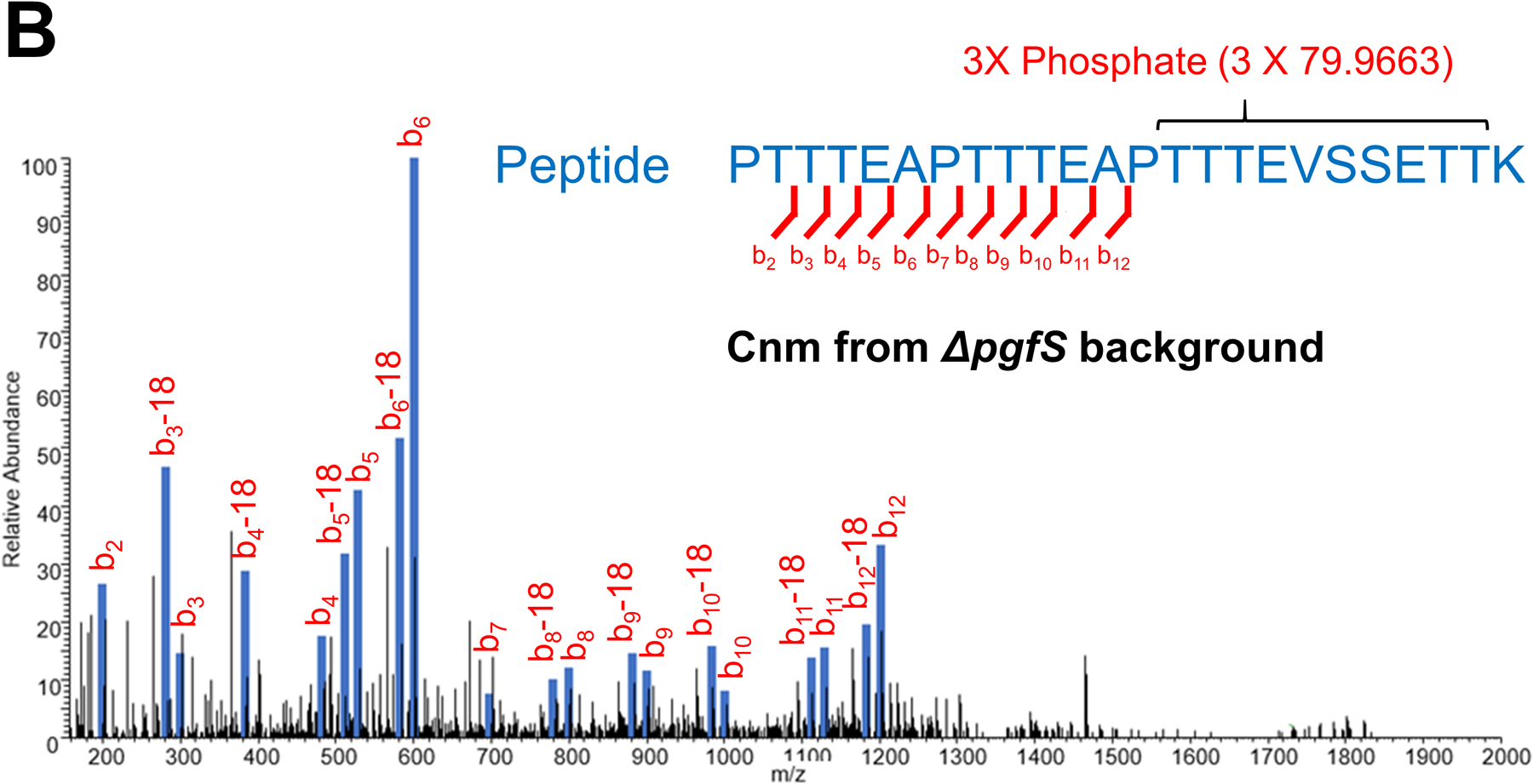
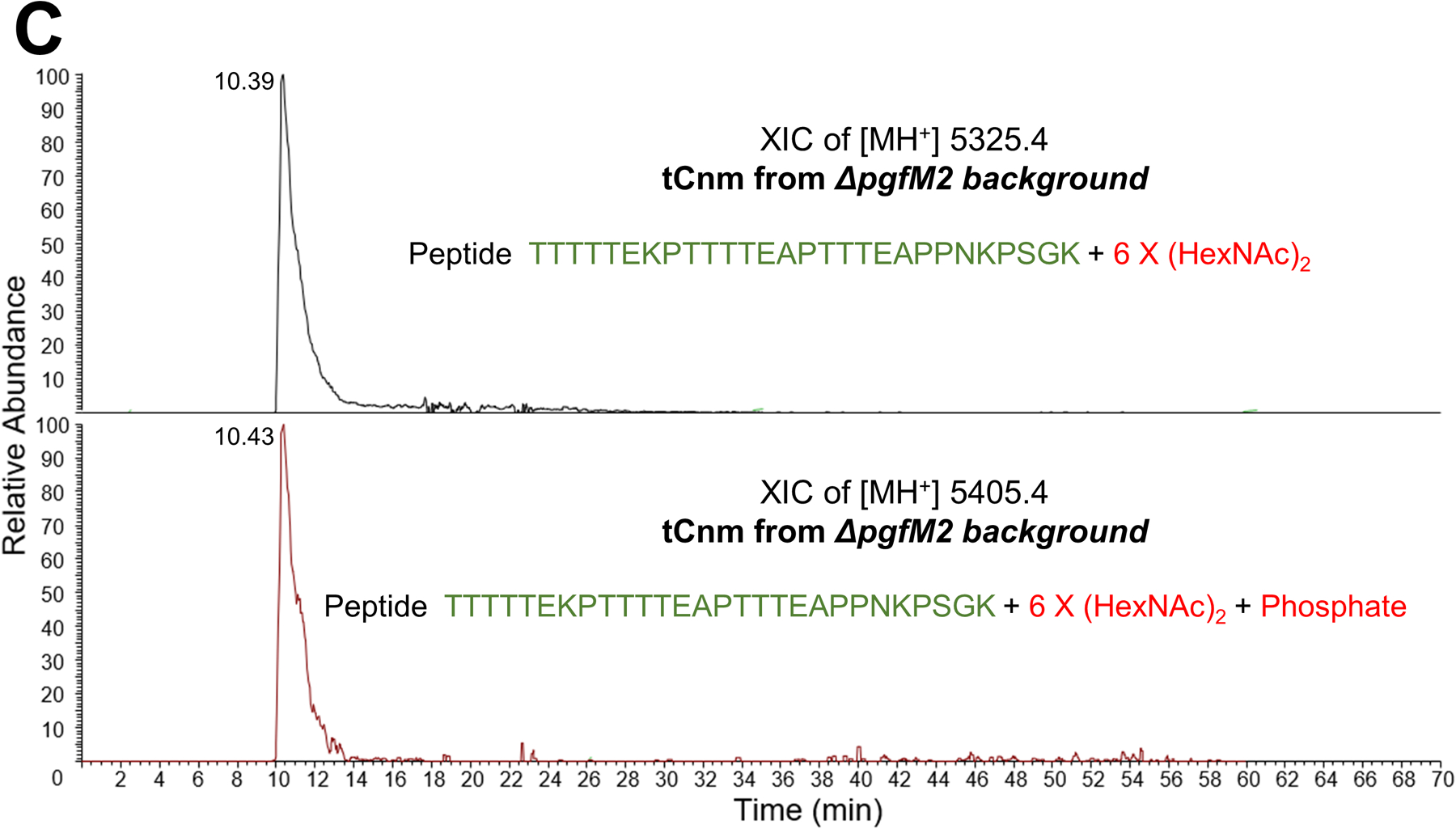
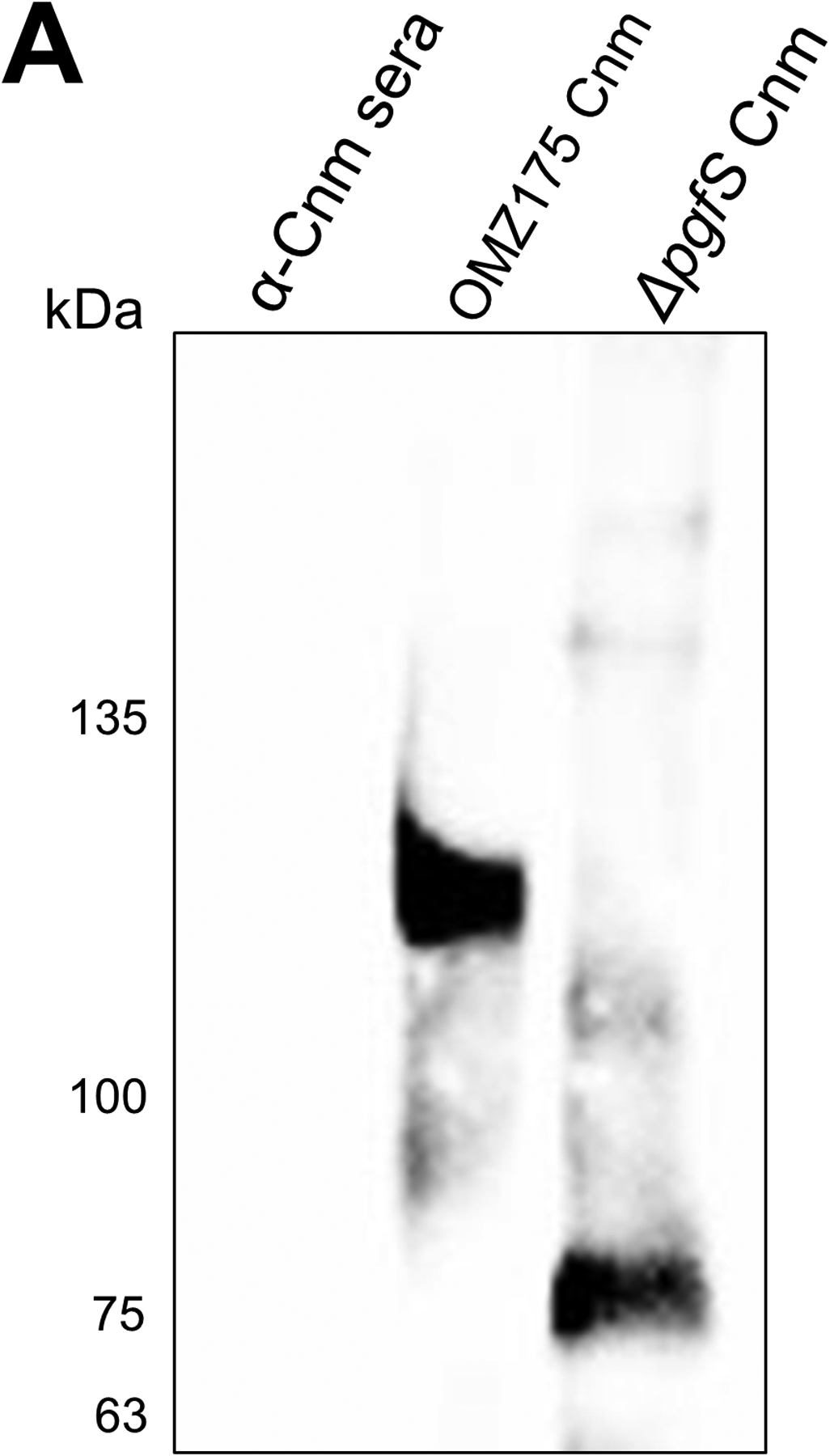
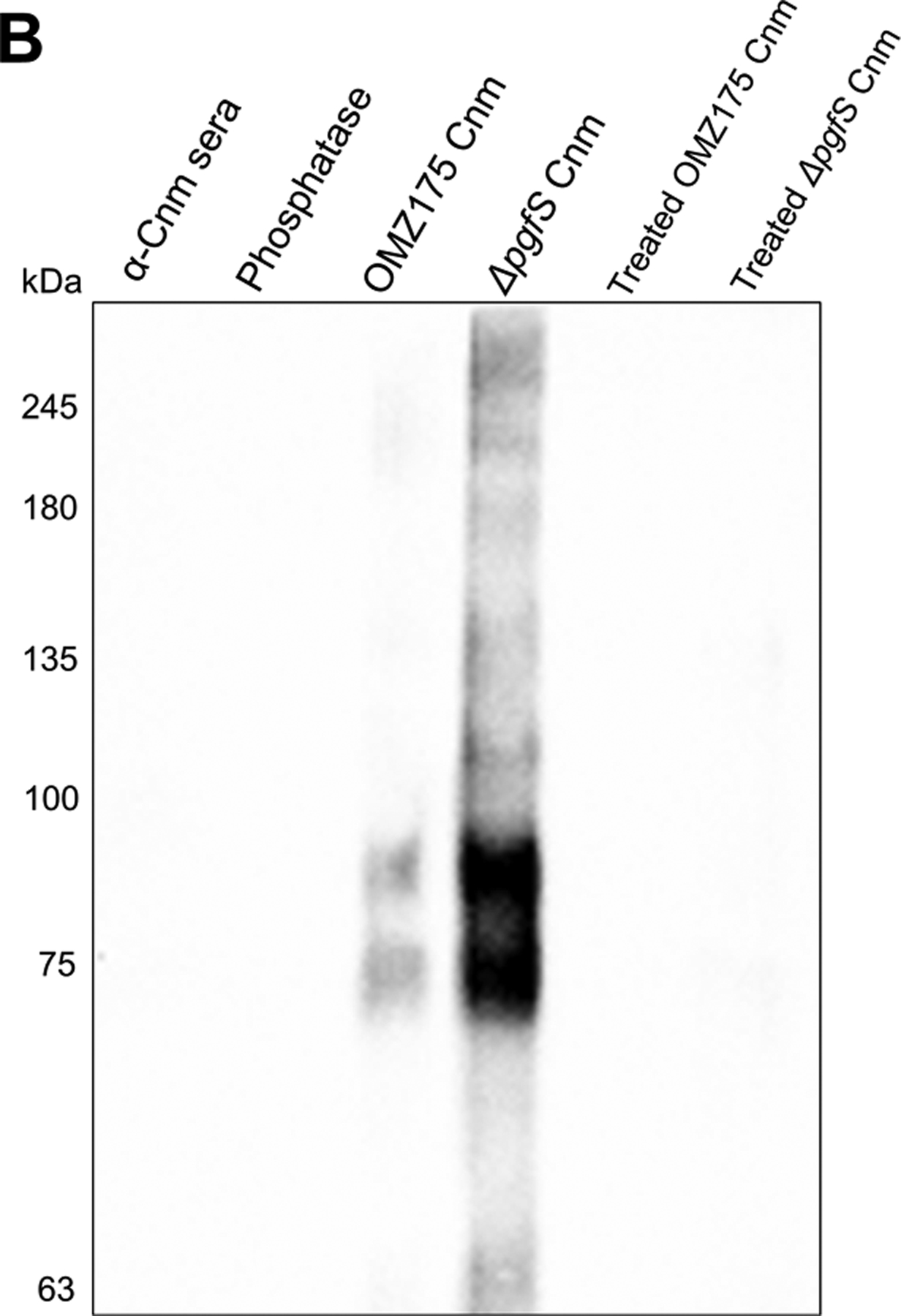
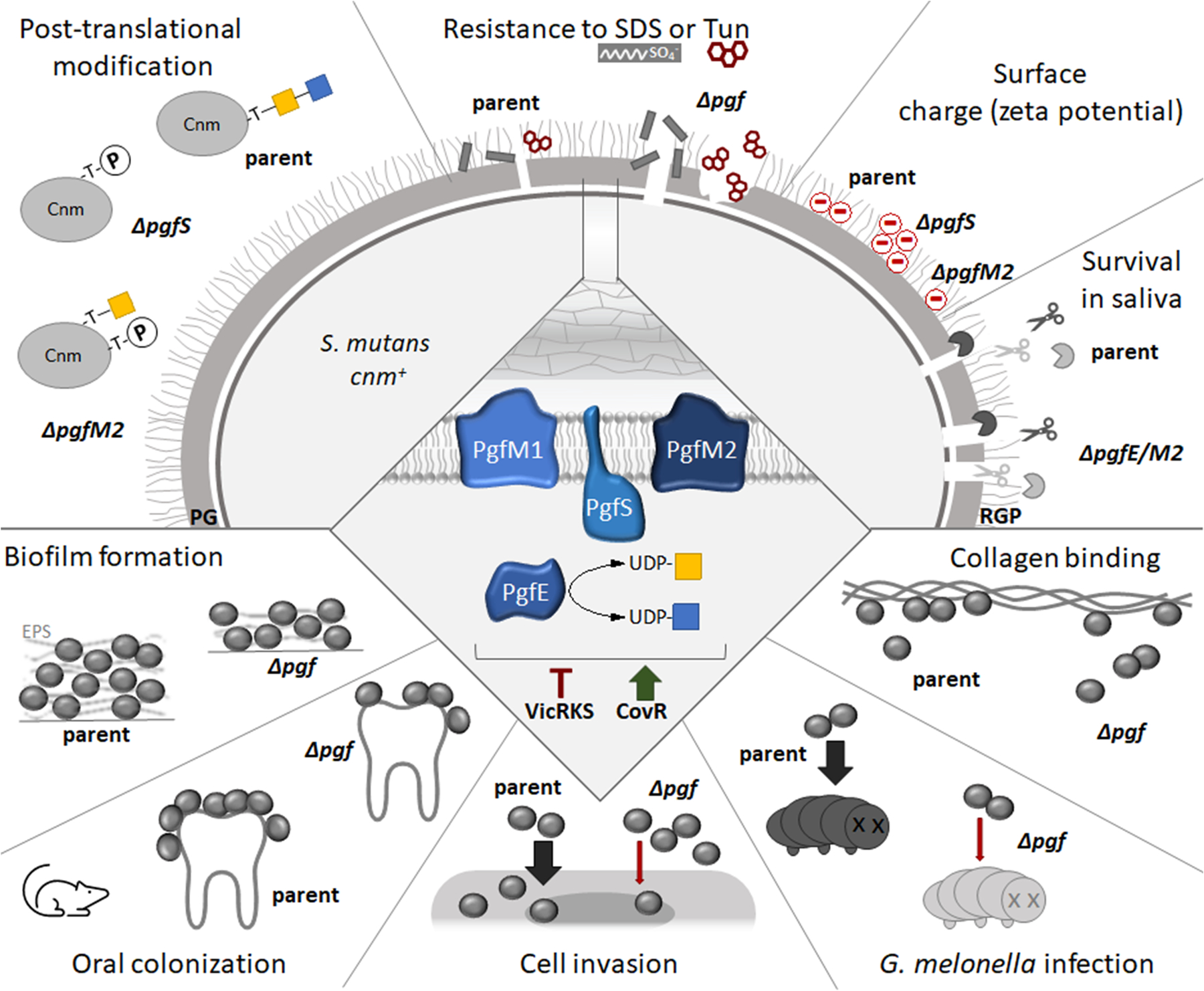
Streptococcus mutans is commonly associated with dental caries and the ability to form biofilms is essential for its pathogenicity. We recently identified the Pgf glycosylation machinery of S. mutans, responsible for the post-translational modification of the surface-associated adhesins Cnm and WapA. Since the four-gene pgf operon (pgfS-pgfM1-pgfE-pgfM2) is part of the S. mutans core genome, we hypothesized that the scope of the Pgf system goes beyond Cnm and WapA glycosylation. In silico analyses and tunicamycin sensitivity assays suggested a functional overlap between the Pgf machinery and the rhamnose-glucose polysaccharide synthesis pathway. Phenotypic characterization of pgf mutants (ΔpgfS, ΔpgfE, ΔpgfM1, ΔpgfM2, and Δpgf) revealed that the Pgf system is important for biofilm formation, surface charge, membrane stability, and survival in human saliva. Moreover, deletion of the entire pgf operon (Δpgf strain) resulted in significantly impaired colonization in a rat oral colonization model. Using Cnm as a model, we showed that Cnm is heavily modified with N-acetyl hexosamines but it becomes heavily phosphorylated with the inactivation of the PgfS glycosyltransferase, suggesting a crosstalk between these two post-translational modification mechanisms. Our results revealed that the Pgf machinery contributes to multiple aspects of S. mutans pathobiology that may go beyond Cnm and WapA glycosylation.
Keywords: Streptococcus mutans; Cnm; Pgf machinery; oral colonization; protein glycosylation; protein phosphorylation.
© 2023 John Wiley & Sons Ltd.
Conflict of interest statement
Conflicts of Interest
The authors declare no conflicts of interest.
Figures

Similar articles
-
Characterization of the pgf operon involved in the posttranslational modification of Streptococcus mutans surface proteins.Sci Rep. 2018 Mar 16;8(1):4705. doi: 10.1038/s41598-018-23170-3. Sci Rep. 2018. PMID: 29549320 Free PMC article.
-
CovR and VicRKX Regulate Transcription of the Collagen Binding Protein Cnm of Streptococcus mutans.J Bacteriol. 2018 Nov 6;200(23):e00141-18. doi: 10.1128/JB.00141-18. Print 2018 Dec 1. J Bacteriol. 2018. PMID: 30201780 Free PMC article.
-
Involvement of the Streptococcus mutans PgfE and GalE 4-epimerases in protein glycosylation, carbon metabolism, and cell division.Glycobiology. 2023 Apr 19;33(3):245-259. doi: 10.1093/glycob/cwad004. Glycobiology. 2023. PMID: 36637425 Free PMC article.
-
Multifaceted roles of Candida albicans and Streptococcus mutans in contributing to polybiofilm infections in early childhood caries.Front Cell Infect Microbiol. 2025 Jun 27;15:1625103. doi: 10.3389/fcimb.2025.1625103. eCollection 2025. Front Cell Infect Microbiol. 2025. PMID: 40654570 Free PMC article. Review.
-
Chlorhexidine treatment for the prevention of dental caries in children and adolescents.Cochrane Database Syst Rev. 2015 Apr 13;2015(4):CD008457. doi: 10.1002/14651858.CD008457.pub2. Cochrane Database Syst Rev. 2015. PMID: 25867816 Free PMC article.
Cited by
-
Glycosylation of serine/threonine-rich intrinsically disordered regions of membrane-associated proteins in streptococci.bioRxiv [Preprint]. 2025 Mar 17:2024.05.05.592596. doi: 10.1101/2024.05.05.592596. bioRxiv. 2025. Update in: Nat Commun. 2025 Apr 29;16(1):4011. doi: 10.1038/s41467-025-58692-8. PMID: 38746434 Free PMC article. Updated. Preprint.
-
Glycosylation of oral bacteria in modulating adhesion and biofilm formation.J Oral Microbiol. 2025 Apr 8;17(1):2486650. doi: 10.1080/20002297.2025.2486650. eCollection 2025. J Oral Microbiol. 2025. PMID: 40213769 Free PMC article. Review.
-
Glycosylation of serine/threonine-rich intrinsically disordered regions of membrane-associated proteins in streptococci.Nat Commun. 2025 Apr 29;16(1):4011. doi: 10.1038/s41467-025-58692-8. Nat Commun. 2025. PMID: 40301326 Free PMC article.
-
Post-translational modifications via serine/threonine phosphorylation and GpsB in Streptococcus mutans.bioRxiv [Preprint]. 2025 Jul 25:2025.07.25.666849. doi: 10.1101/2025.07.25.666849. bioRxiv. 2025. PMID: 40777345 Free PMC article. Preprint.
References
-
- Andresen S, Fantone K, Chapla D, Rada B, Moremen KW, Pierce M, and Szymanski CM (2022) Human Intelectin-1 Promotes Cellular Attachment and Neutrophil Killing of Streptococcus pneumoniae in a Serotype-Dependent Manner. Infection and Immunity 90 https://journals.asm.org/doi/10.1128/iai.00682-21. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T90 DE021990/DE/NIDCR NIH HHS/United States
- R90 DE022530/DE/NIDCR NIH HHS/United States
- T32 GM107004/GM/NIGMS NIH HHS/United States
- R01 DE022559/DE/NIDCR NIH HHS/United States
- R21 AI147097/AI/NIAID NIH HHS/United States
- DE022559/DE/NIDCR NIH HHS/United States
- R01 HL136707/HL/NHLBI NIH HHS/United States
- R24 GM137782/GM/NIGMS NIH HHS/United States
- DE021990-11/DE/NIDCR NIH HHS/United States
- DE022559/DE/NIDCR NIH HHS/United States
- DE021990-11/DE/NIDCR NIH HHS/United States
- NIGMS Training grant T32GM107004
- R24GM137782/University of Georgia's Complex Carbohydrate Research Center, National Institutes of Health (NIH)-funded R24 grant
LinkOut - more resources
Full Text Sources
