

Mechanism of target site selection by type V-K CRISPR-associated transposases
- PMID: 37972161
- PMCID: PMC10771339
- DOI: 10.1126/science.adj8543
Mechanism of target site selection by type V-K CRISPR-associated transposases
Abstract
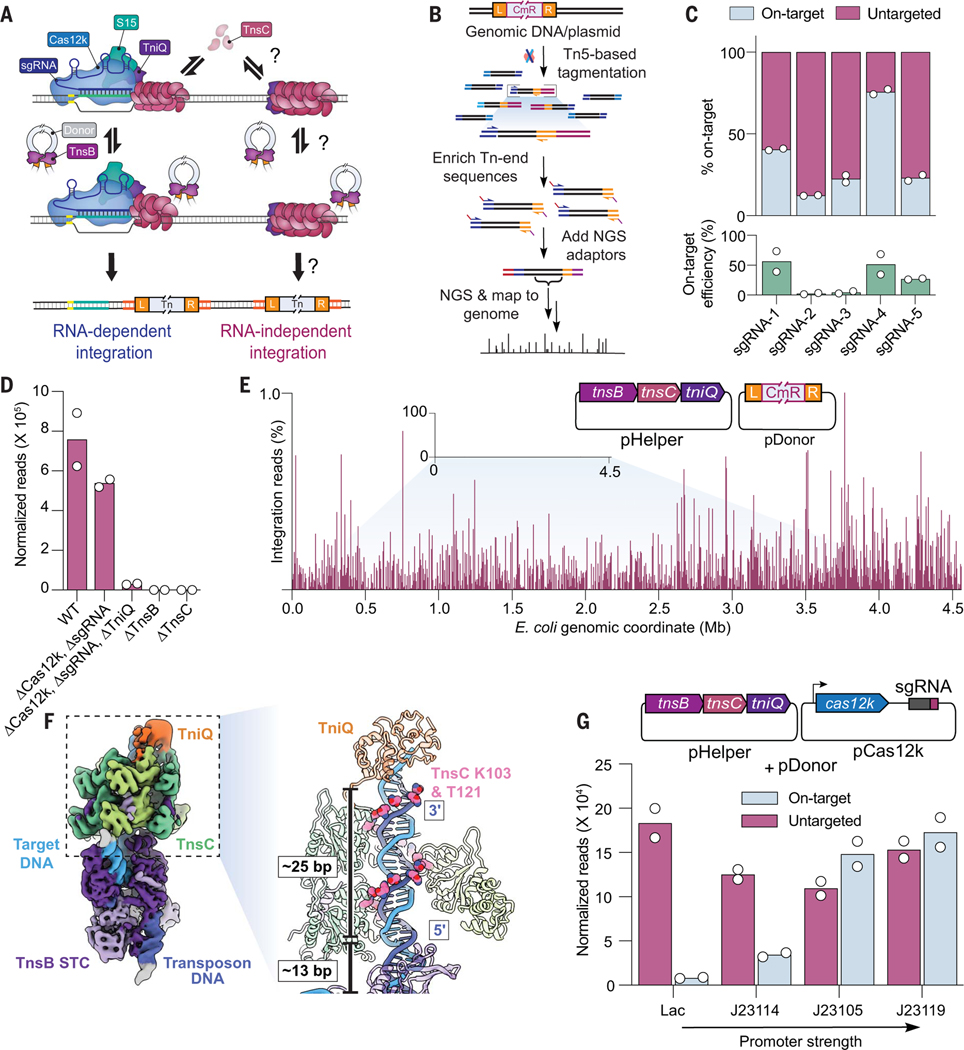
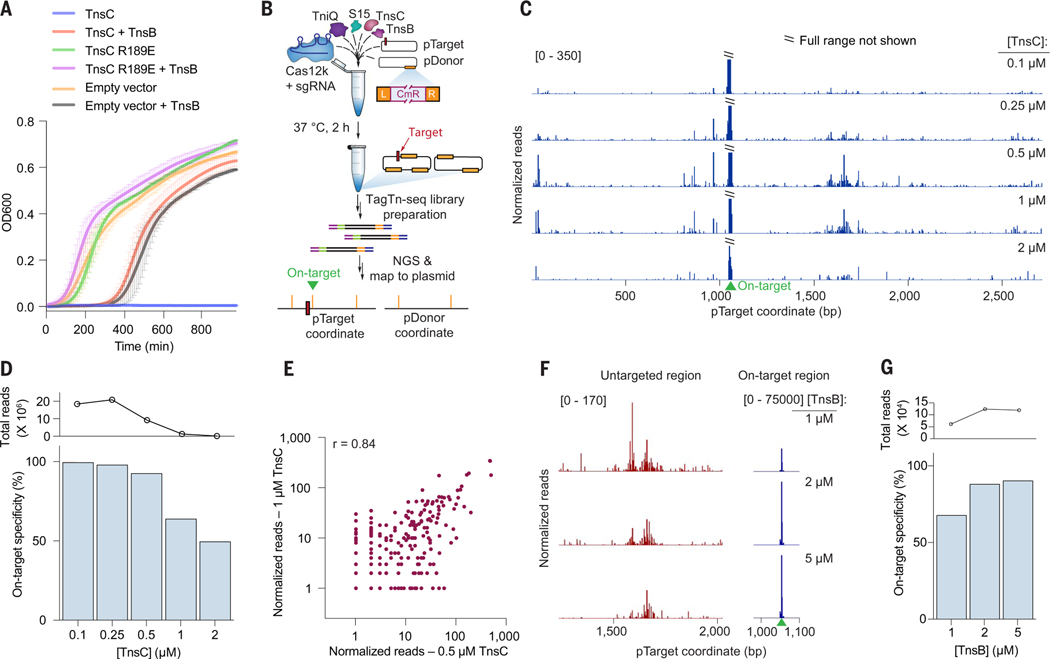
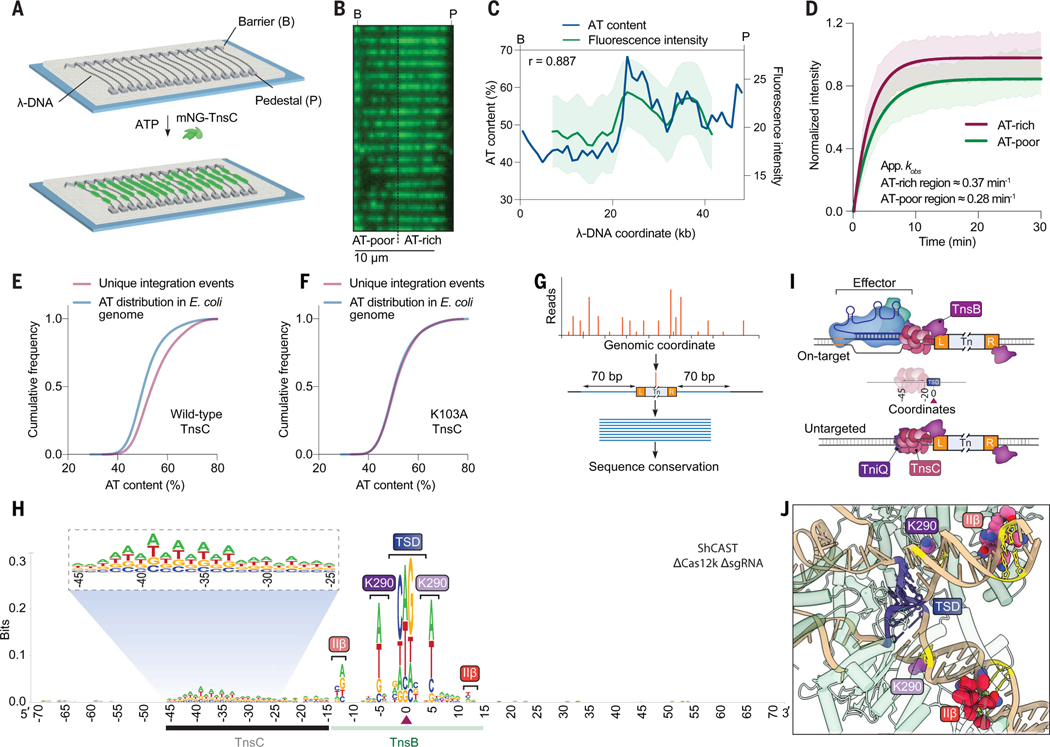
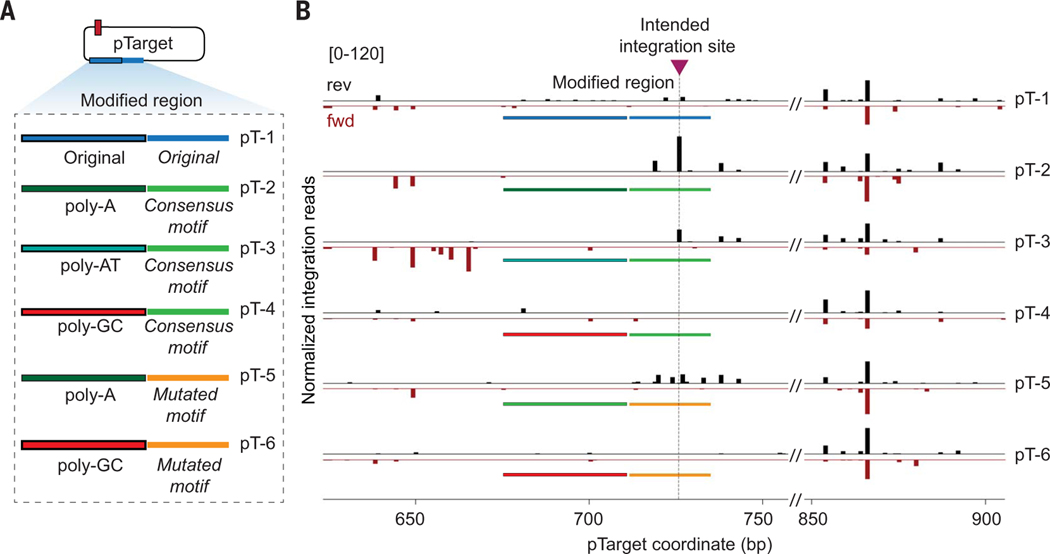
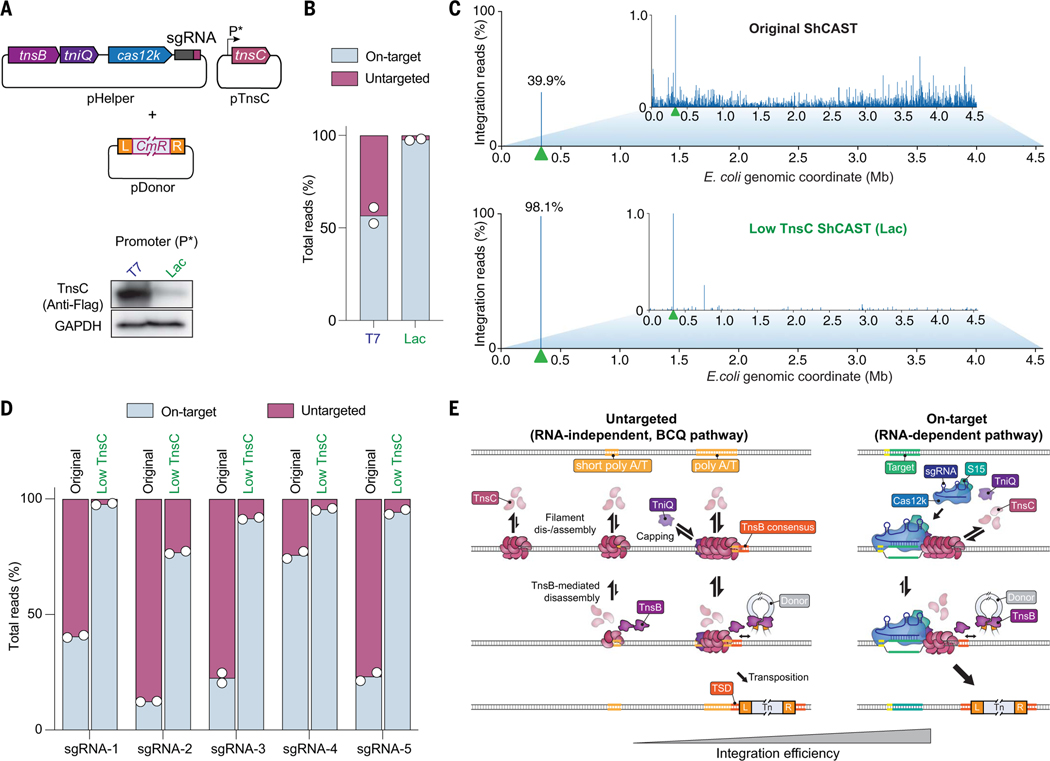
CRISPR-associated transposases (CASTs) repurpose nuclease-deficient CRISPR effectors to catalyze RNA-guided transposition of large genetic payloads. Type V-K CASTs offer potential technology advantages but lack accuracy, and the molecular basis for this drawback has remained elusive. Here, we reveal that type V-K CASTs maintain an RNA-independent, "untargeted" transposition pathway alongside RNA-dependent integration, driven by the local availability of TnsC filaments. Using cryo-electron microscopy, single-molecule experiments, and high-throughput sequencing, we found that a minimal, CRISPR-less transpososome preferentially directs untargeted integration at AT-rich sites, with additional local specificity imparted by TnsB. By exploiting this knowledge, we suppressed untargeted transposition and increased type V-K CAST specificity up to 98.1% in cells without compromising on-target integration efficiency. These findings will inform further engineering of CAST systems for accurate, kilobase-scale genome engineering applications.
Conflict of interest statement
Figures
Update of
-
Mechanism of target site selection by type V-K CRISPR-associated transposases.bioRxiv [Preprint]. 2023 Jul 14:2023.07.14.548620. doi: 10.1101/2023.07.14.548620. bioRxiv. 2023. Update in: Science. 2023 Nov 17;382(6672):eadj8543. doi: 10.1126/science.adj8543. PMID: 37503092 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
