

A toolbox of astrocyte-specific, serotype-independent adeno-associated viral vectors using microRNA targeting sequences
- PMID: 37973910
- PMCID: PMC10654773
- DOI: 10.1038/s41467-023-42746-w
A toolbox of astrocyte-specific, serotype-independent adeno-associated viral vectors using microRNA targeting sequences
Abstract
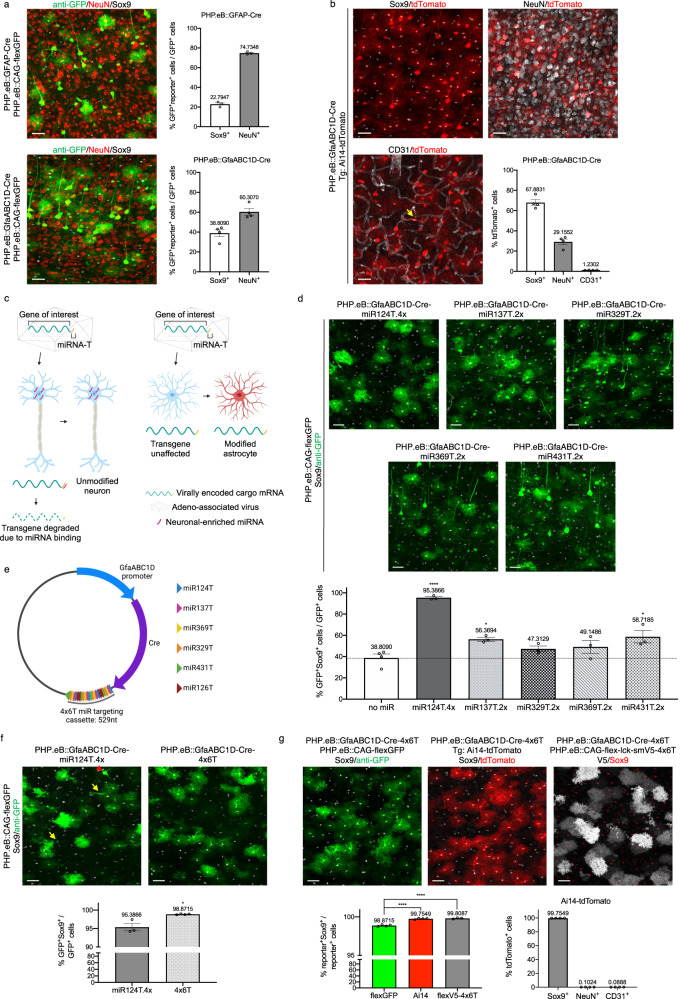
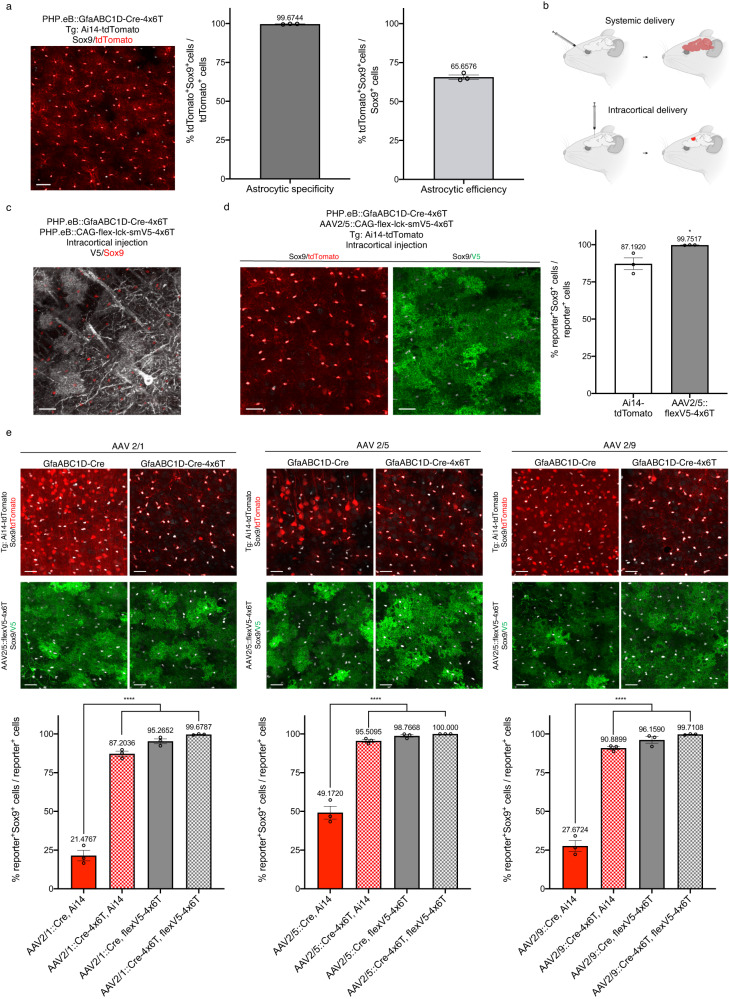
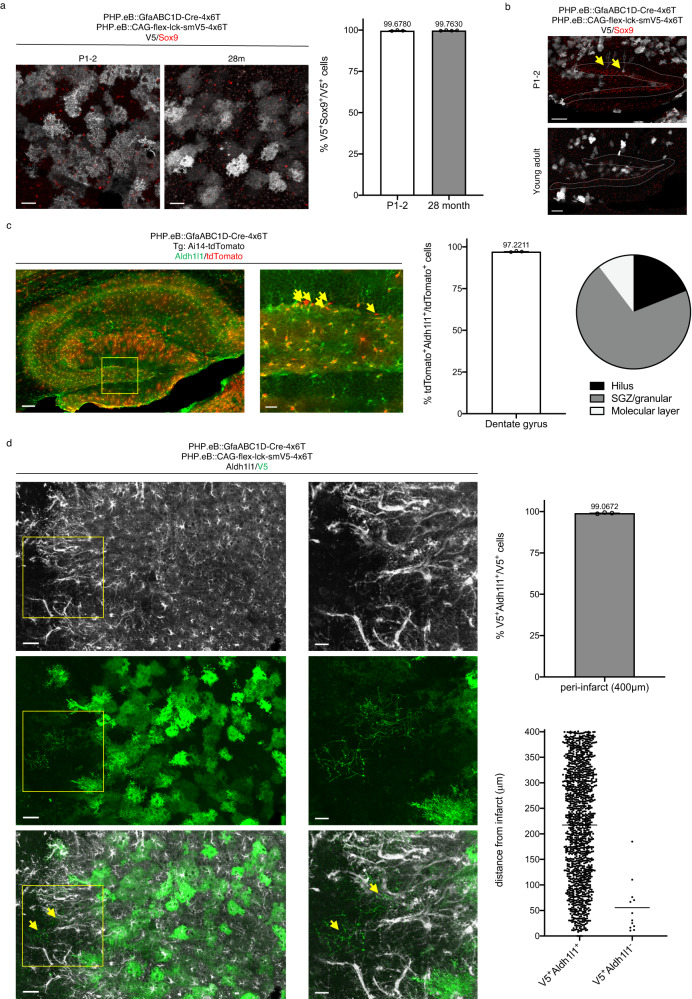
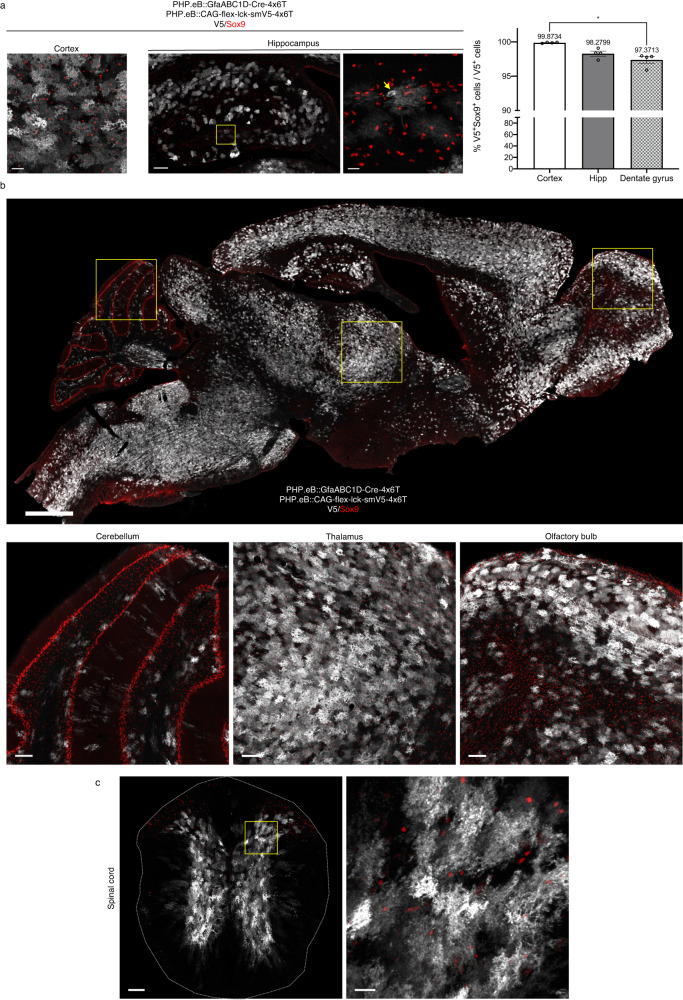
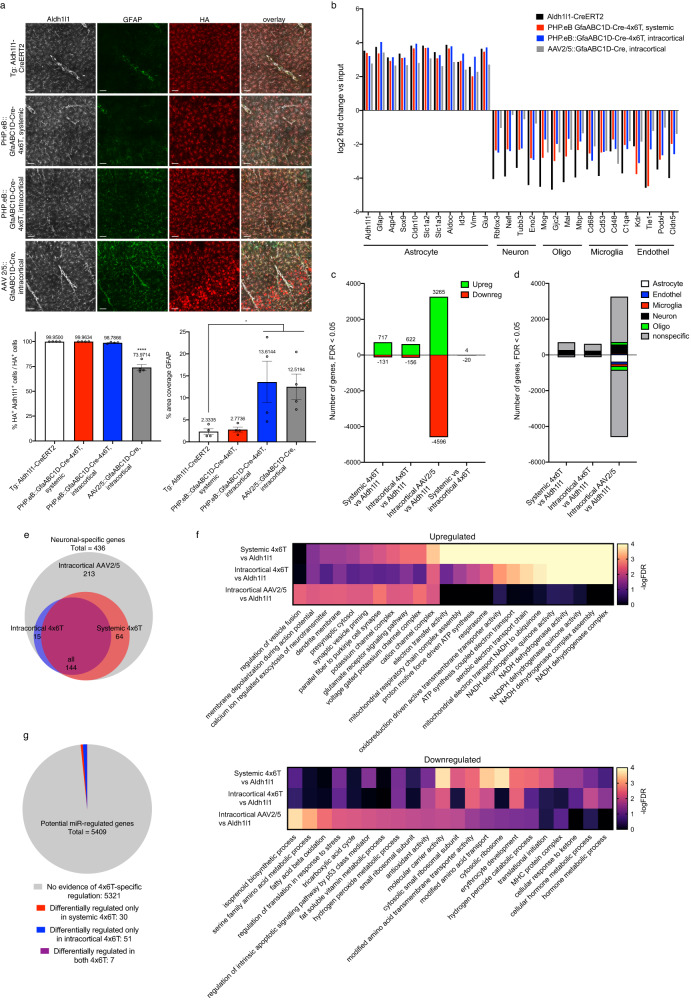
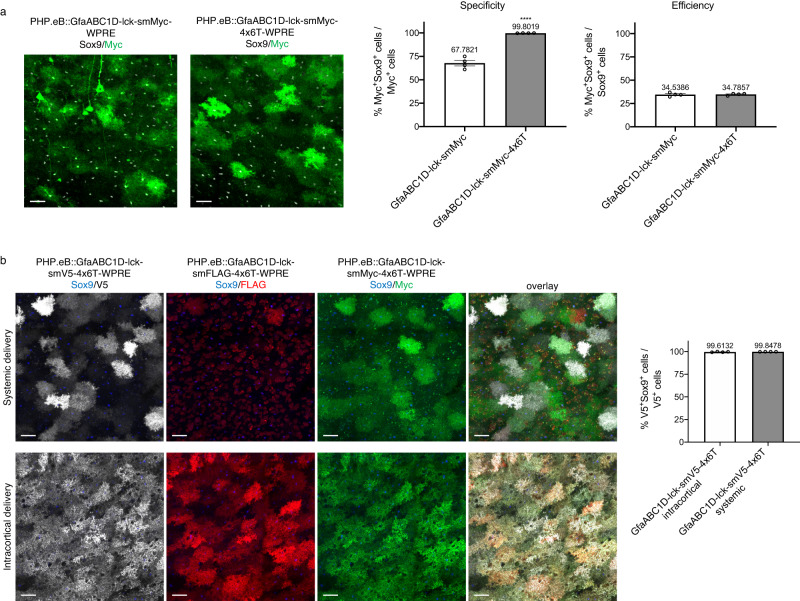
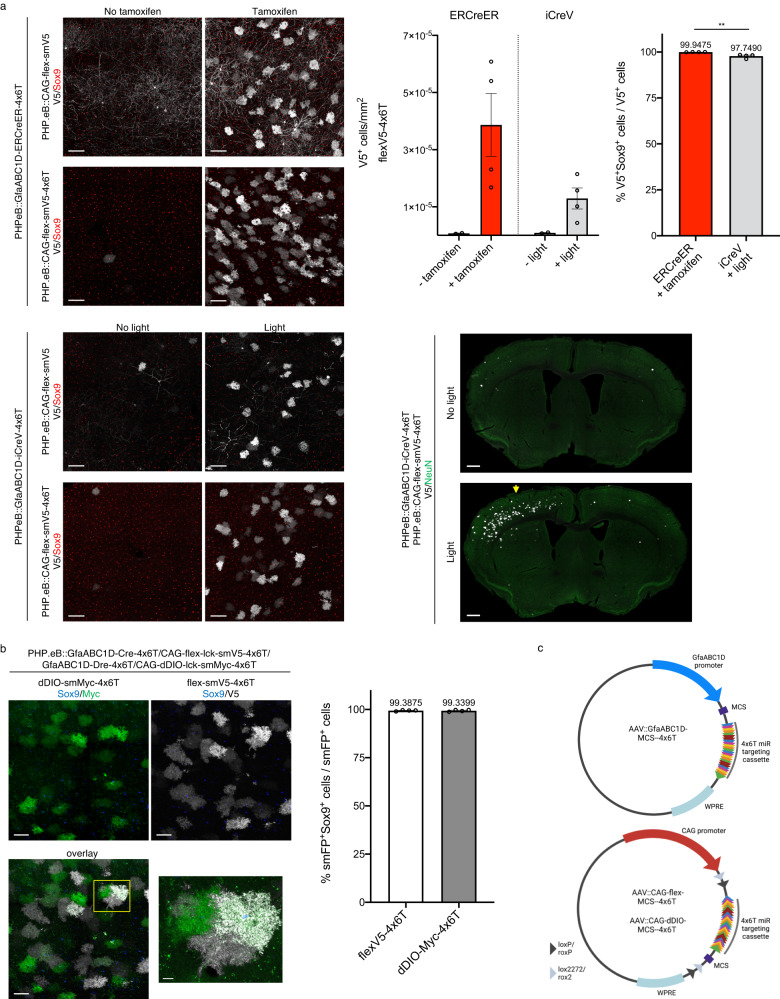
Astrocytes, one of the most prevalent cell types in the central nervous system (CNS), are critically involved in neural function. Genetically manipulating astrocytes is an essential tool in understanding and affecting their roles. Adeno-associated viruses (AAVs) enable rapid genetic manipulation; however, astrocyte specificity of AAVs can be limited, with high off-target expression in neurons and sparsely in endothelial cells. Here, we report the development of a cassette of four copies of six miRNA targeting sequences (4x6T) which triggers transgene degradation specifically in neurons and endothelial cells. In combination with the GfaABC1D promoter, 4x6T increases astrocytic specificity of Cre with a viral reporter from <50% to >99% in multiple serotypes in mice, and confers astrocyte specificity in multiple recombinases and reporters. We also present empty vectors to add 4x6T to other cargo, independently and in Cre/Dre-dependent forms. This toolbox of AAVs allows rapid manipulation of astrocytes throughout the CNS, is compatible with different AAV serotypes, and demonstrates the efficacy of using multiplexed miRNA targeting sequences to decrease expression in multiple off-target cell populations simultaneously.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Cellular selectivity of AAV serotypes for gene delivery in neurons and astrocytes by neonatal intracerebroventricular injection.PLoS One. 2017 Dec 15;12(12):e0188830. doi: 10.1371/journal.pone.0188830. eCollection 2017. PLoS One. 2017. PMID: 29244806 Free PMC article.
-
Systemic application of AAV vectors targeting GFAP-expressing astrocytes in Z-Q175-KI Huntington's disease mice.Mol Cell Neurosci. 2016 Dec;77:76-86. doi: 10.1016/j.mcn.2016.10.007. Epub 2016 Oct 27. Mol Cell Neurosci. 2016. PMID: 27989734
-
tTARGIT AAVs mediate the sensitive and flexible manipulation of intersectional neuronal populations in mice.Elife. 2021 Mar 11;10:e66835. doi: 10.7554/eLife.66835. Elife. 2021. PMID: 33704065 Free PMC article.
-
Genetic Constructs for the Control of Astrocytes' Activity.Cells. 2021 Jun 25;10(7):1600. doi: 10.3390/cells10071600. Cells. 2021. PMID: 34202359 Free PMC article. Review.
-
Cre Activated and Inactivated Recombinant Adeno-Associated Viral Vectors for Neuronal Anatomical Tracing or Activity Manipulation.Curr Protoc Neurosci. 2015 Jul 1;72:1.24.1-1.24.15. doi: 10.1002/0471142301.ns0124s72. Curr Protoc Neurosci. 2015. PMID: 26131660 Free PMC article. Review.
Cited by
-
GEARBOCS: An Adeno Associated Virus Tool for In Vivo Gene Editing in Astrocytes.bioRxiv [Preprint]. 2024 Oct 10:2023.01.17.524433. doi: 10.1101/2023.01.17.524433. bioRxiv. 2024. PMID: 36711516 Free PMC article. Preprint.
-
Astrocyte-Mediated Neuroinflammation in Neurological Conditions.Biomolecules. 2024 Sep 25;14(10):1204. doi: 10.3390/biom14101204. Biomolecules. 2024. PMID: 39456137 Free PMC article. Review.
-
Efficient gene delivery admitted by small metabolites specifically targeting astrocytes in the mouse brain.Mol Ther. 2025 Mar 5;33(3):1166-1179. doi: 10.1016/j.ymthe.2025.01.006. Epub 2025 Jan 10. Mol Ther. 2025. PMID: 39799395
-
Specific targeting of brain endothelial cells using enhancer AAV vectors.Neuron. 2025 May 21;113(10):1562-1578.e6. doi: 10.1016/j.neuron.2025.03.031. Neuron. 2025. PMID: 40403707
-
Preclinical Insights into the Role of Kir4.1 in Chronic Pain and Depression: Mechanisms and Therapeutic Potential.Biomolecules. 2025 Jan 23;15(2):165. doi: 10.3390/biom15020165. Biomolecules. 2025. PMID: 40001468 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
