

Pericytes are protective in experimental pneumococcal meningitis through regulating leukocyte infiltration and blood-brain barrier function
- PMID: 37978545
- PMCID: PMC10655320
- DOI: 10.1186/s12974-023-02938-z
Pericytes are protective in experimental pneumococcal meningitis through regulating leukocyte infiltration and blood-brain barrier function
Abstract
Background: Brain pericytes participate in the regulation of cerebral blood flow and the maintenance of blood-brain barrier integrity. Because of their perivascular localization, their receptor repertoire, and their potential ability to respond to inflammatory and infectious stimuli by producing various cytokines and chemokines, these cells are also thought to play an active role in the immune response to brain infections. This assumption is mainly supported by in vitro studies, investigations in in vivo disease models are largely missing. Here, we analysed the role of brain pericytes in pneumococcal meningitis, in vitro and in vivo in two animal models of pneumococcal meningitis.
Methods: Primary murine and human pericytes were stimulated with increasing concentrations of different serotypes of Streptococcus pneumoniae in the presence or absence of Toll-like receptor inhibitors and their cell viability and cytokine production were monitored. To gain insight into the role of pericytes in brain infection in vivo, we performed studies in a zebrafish embryo model of pneumococcal meningitis in which pericytes were pharmacologically depleted. Furthermore, we analyzed the impact of genetically induced pericyte ablation on disease progression, intracranial complications, and brain inflammation in an adult mouse model of this disease.
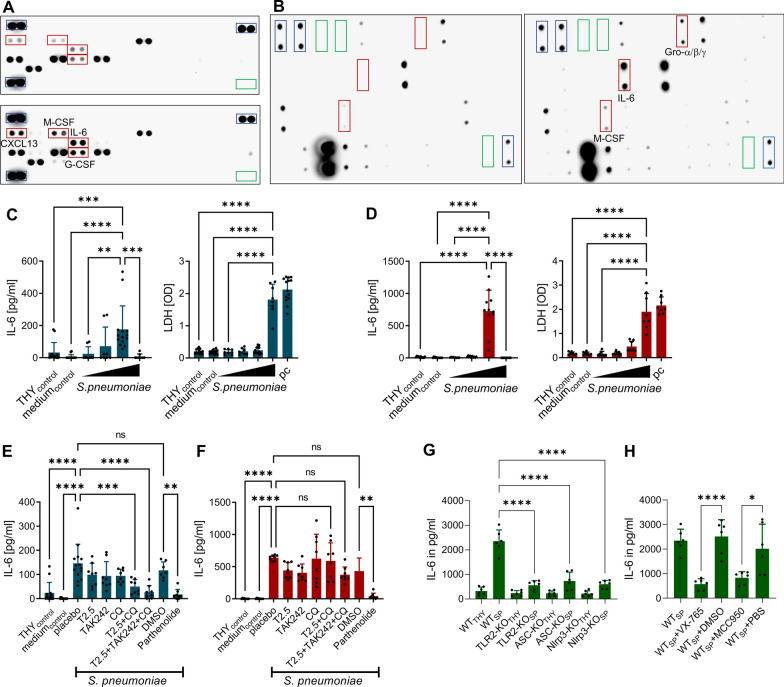
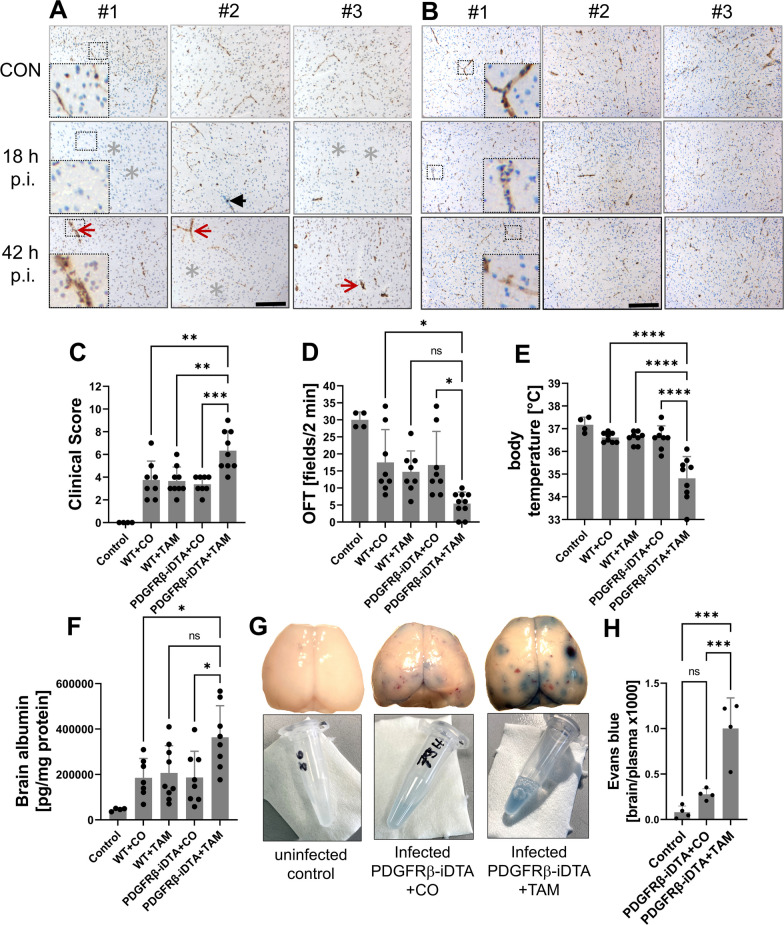
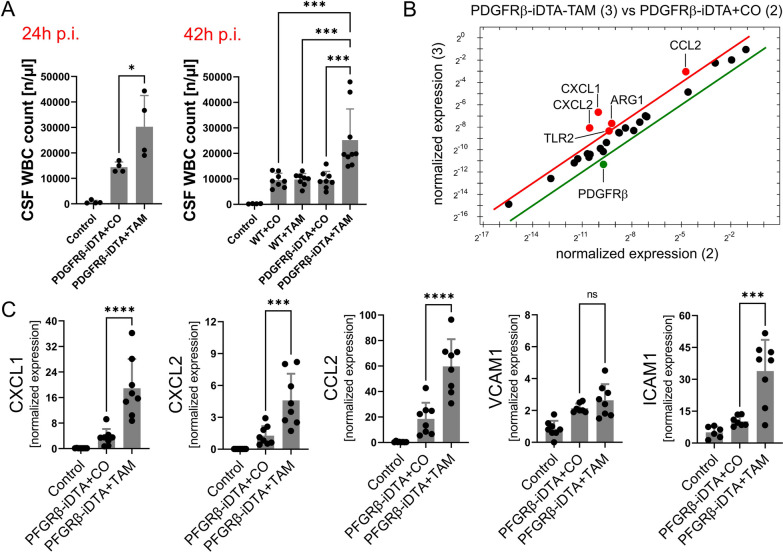
Results: Both murine and human pericytes reacted to pneumococcal exposure with the release of selected cytokines. This cytokine release is pneumolysin-dependent, TLR-dependent in murine (but not human) pericytes and can be significantly increased by macrophage-derived IL-1b. Pharmacological depletion of pericytes in zebrafish embryos resulted in increased cerebral edema and mortality due to pneumococcal meningitis. Correspondingly, in an adult mouse meningitis model, a more pronounced blood-brain barrier disruption and leukocyte infiltration, resulting in an unfavorable disease course, was observed following genetic pericyte ablation. The degree of leukocyte infiltration positively correlated with an upregulation of chemokine expression in the brains of pericyte-depleted mice.
Conclusions: Our findings show that pericytes play a protective role in pneumococcal meningitis by impeding leukocyte migration and preventing blood-brain barrier breaching. Thus, preserving the integrity of the pericyte population has the potential as a new therapeutic strategy in pneumococcal meningitis.
Keywords: Blood–brain barrier; Pericytes; Pneumococcal meningitis; Streptococcus pneumoniae.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Pinho-Ribeiro FA, Deng L, Neel DV, Erdogan O, Basu H, Yang D, Choi S, Walker AJ, Carneiro-Nascimento S, He K, Wu G, Stevens B, Doran KS, Levy D, Chiu IM. Bacteria hijack a meningeal neuroimmune axis to facilitate brain invasion. Nature. 2023;615:472. doi: 10.1038/s41586-023-05753-x. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
