

Lactobacillus Intestinalis Primes Epithelial Cells to Suppress Colitis-Related Th17 Response by Host-Microbe Retinoic Acid Biosynthesis
- PMID: 37983567
- PMCID: PMC10754072
- DOI: 10.1002/advs.202303457
Lactobacillus Intestinalis Primes Epithelial Cells to Suppress Colitis-Related Th17 Response by Host-Microbe Retinoic Acid Biosynthesis
Abstract
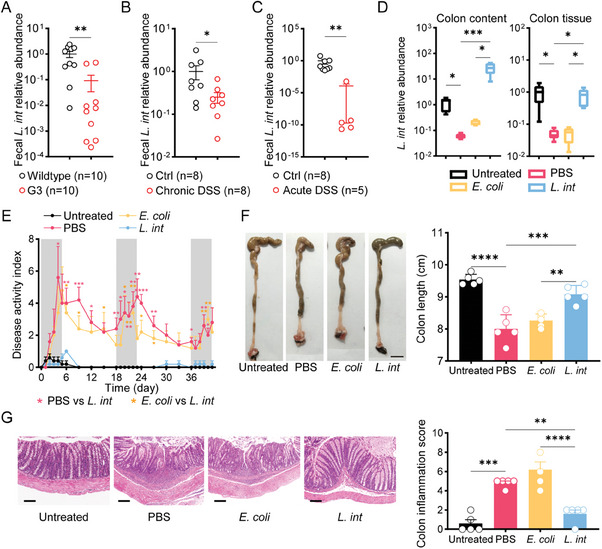
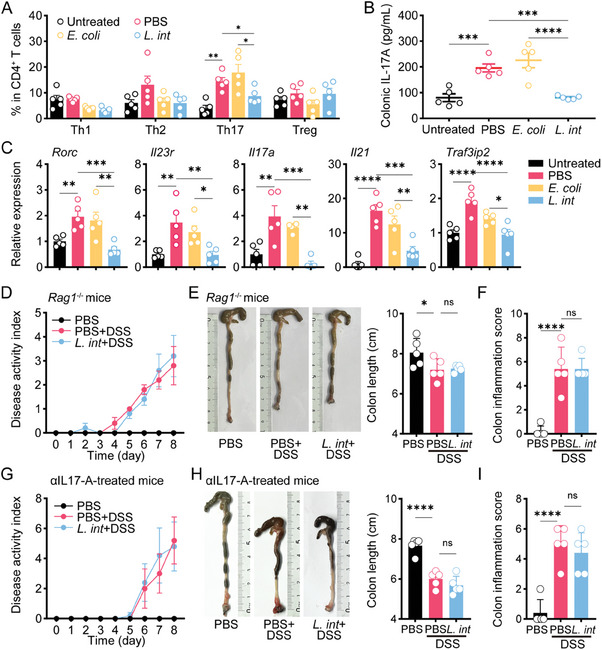
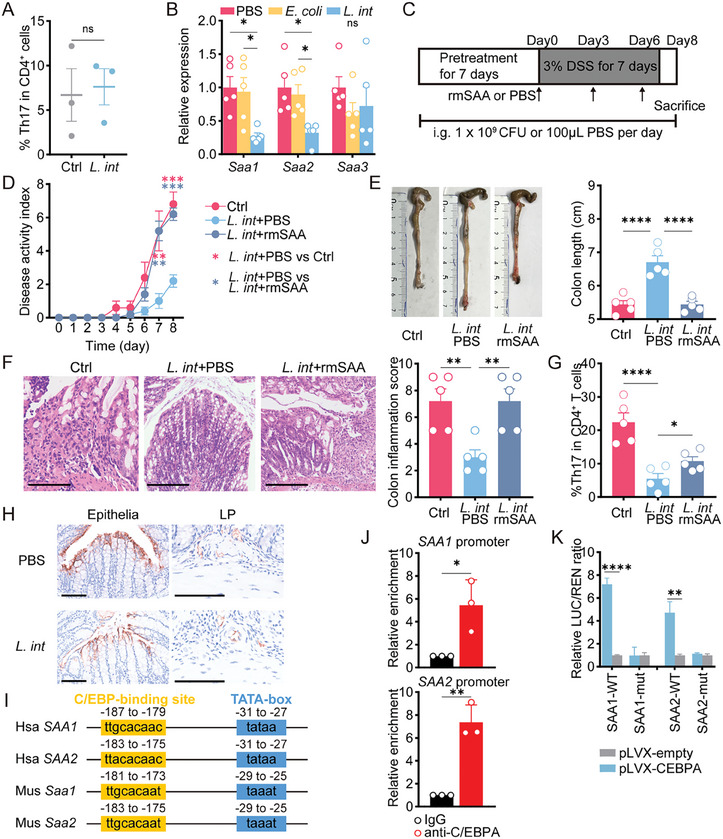
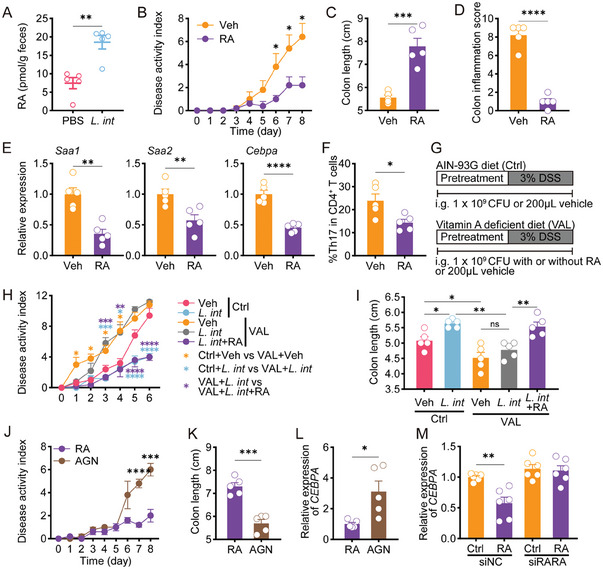
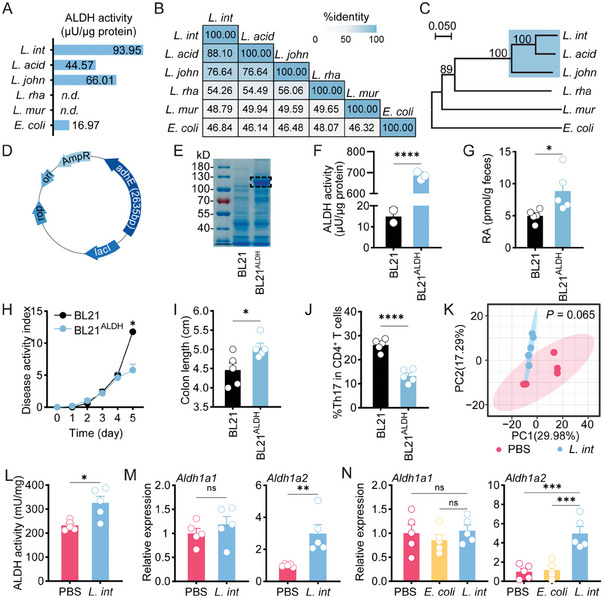
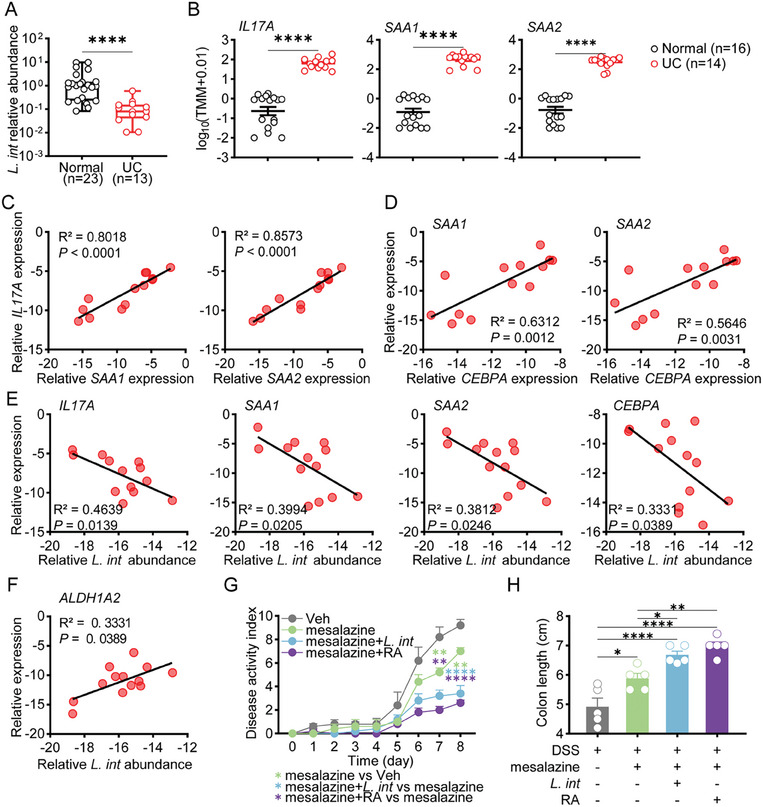
Gut microbiome is integral to the pathogenesis of ulcerative colitis. A novel probiotic Lactobacillus intestinalis (L. intestinalis) exerts a protective effect against dextran sodium sulfate-induced colitis in mice. Based on flow cytometry, colitis-associated Th17 cells are the target of L. intestinalis, which is supported by the lack of protective effects of L. intestinalis in T cell-null Rag1-/- mice or upon anti-IL-17-A antibody-treated mice. Although L. intestinalis exerts no direct effect on T cell differentiation, it decreases C/EBPA-driven gut epithelial SAA1 and SAA2 production, which in turn impairs Th17 cell differentiation. Cometabolism of L. intestinalis ALDH and host ALDH1A2 contributed to elevated biosynthesis of retinoic acid (RA), which accounts for the anti-colitis effect in RAR-α -mediated way. In a cohort of ulcerative colitis patients, it is observed that fecal abundance of L. intestinalis is negatively associated with the C/EBPA-SAA1/2-Th17 axis. Finally, L. intestinalis has a synergistic effect with mesalazine in alleviating murine colitis. In conclusion, L. intestinalis and associated metabolites, RA, have potential therapeutic effects for suppressing colonic inflammation by modulating the crosstalk between intestinal epithelia and immunity.
Keywords: Th17; colitis; microbes; retinoic acid.
© 2023 The Authors. Advanced Science published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous