

H. pylori-induced NF-κB-PIEZO1-YAP1-CTGF axis drives gastric cancer progression and cancer-associated fibroblast-mediated tumour microenvironment remodelling
- PMID: 37983931
- PMCID: PMC10659770
- DOI: 10.1002/ctm2.1481
H. pylori-induced NF-κB-PIEZO1-YAP1-CTGF axis drives gastric cancer progression and cancer-associated fibroblast-mediated tumour microenvironment remodelling
Abstract
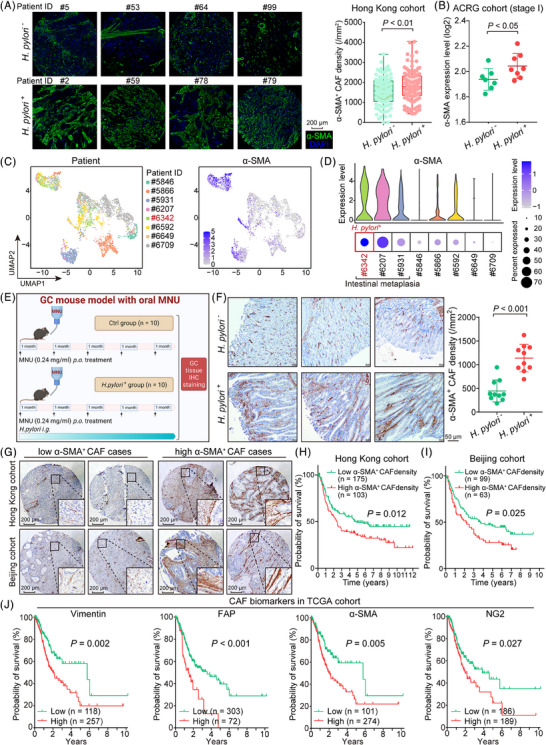
Background: Gastric cancer (GC) is one of the most common tumours in East Asia countries and is associated with Helicobacter pylori infection. H. pylori utilizes virulence factors, CagA and VacA, to up-regulate pro-inflammatory cytokines and activate NF-κB signaling. Meanwhile, the PIEZO1 upregulation and cancer-associated fibroblast (CAF) enrichment were found in GC progression. However, the mechanisms of PIEZO1 upregulation and its involvement in GC progression have not been fully elucidated.
Methods: The CAF enrichment and clinical significance were investigated in animal models and primary samples. The expression of NF-κB and PIEZO1 in GC was confirmed by immunohistochemistry staining, and expression correlation was analysed in multiple GC datasets. GSEA and Western blot analysis revealed the YAP1-CTGF axis regulation by PIEZO1. The stimulatory effects of CTGF on CAFs were validated by the co-culture system and animal studies. Patient-derived organoid and peritoneal dissemination models were employed to confirm the role of the PIEZO1-YAP1-CTGF cascade in GC.
Results: Both CAF signature and PIEZO1 were positively correlated with H. pylori infection. PIEZO1, a mechanosensor, was confirmed as a direct downstream of NF-κB to promote the transformation from intestinal metaplasia to GC. Mechanistic studies revealed that PIEZO1 transduced the oncogenic signal from NF-κB into YAP1 signaling, a well-documented oncogenic pathway in GC progression. PIEZO1 expression was positively correlated with the YAP1 signature (CTGF, CYR61, and c-Myc, etc.) in primary samples. The secreted CTGF by cancer cells stimulated the CAF infiltration to form a stiffened collagen-enrichment microenvironment, thus activating PIEZO1 to form a positive feedback loop. Both PIEZO1 depletion by shRNA and CTGF inhibition by Procyanidin C1 enhanced the efficacy of 5-FU in suppressing the GC cell peritoneal metastasis.
Conclusion: This study elucidates a novel driving PIEZO1-YAP1-CTGF force, which opens a novel therapeutic avenue to block the transformation from precancerous lesions to GC. H. pylori-NF-κB activates the PIEZO1-YAP1-CTGF axis to remodel the GC microenvironment by promoting CAF infiltration. Targeting PIEZO1-YAP1-CTGF plus chemotherapy might serve as a potential therapeutic option to block GC progression and peritoneal metastasis.
Keywords: CTGF; H. pylori; NF-κB; PIEZO1; YAP1; cancer-associated fibroblast; gastric cancer.
© 2023 The Authors. Clinical and Translational Medicine published by John Wiley & Sons Australia, Ltd on behalf of Shanghai Institute of Clinical Bioinformatics.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous