

The Onset of Molecule-Spanning Dynamics in Heat Shock Protein Hsp90
- PMID: 37984887
- PMCID: PMC10754087
- DOI: 10.1002/advs.202304262
The Onset of Molecule-Spanning Dynamics in Heat Shock Protein Hsp90
Abstract
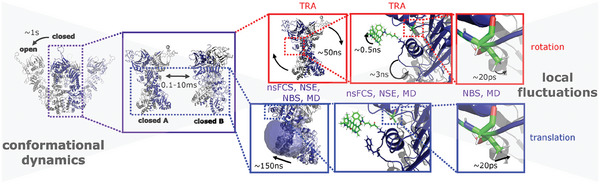
Protein dynamics have been investigated on a wide range of time scales. Nano- and picosecond dynamics have been assigned to local fluctuations, while slower dynamics have been attributed to larger conformational changes. However, it is largely unknown how fast (local) fluctuations can lead to slow global (allosteric) changes. Here, fast molecule-spanning dynamics on the 100 to 200 ns time scale in the heat shock protein 90 (Hsp90) are shown. Global real-space movements are assigned to dynamic modes on this time scale, which is possible by a combination of single-molecule fluorescence, quasi-elastic neutron scattering and all-atom molecular dynamics (MD) simulations. The time scale of these dynamic modes depends on the conformational state of the Hsp90 dimer. In addition, the dynamic modes are affected to various degrees by Sba1, a co-chaperone of Hsp90, depending on the location within Hsp90, which is in very good agreement with MD simulations. Altogether, this data is best described by fast molecule-spanning dynamics, which precede larger conformational changes in Hsp90 and might be the molecular basis for allostery. This integrative approach provides comprehensive insights into molecule-spanning dynamics on the nanosecond time scale for a multi-domain protein.
Keywords: heat shock protein 90; molecular dynamics simulations; neutron scattering; protein dynamics; single molecule fluorescence.
© 2023 The Authors. Advanced Science published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Figures






Similar articles
-
Exploring Mechanisms of Allosteric Regulation and Communication Switching in the Multiprotein Regulatory Complexes of the Hsp90 Chaperone with Cochaperones and Client Proteins: Atomistic Insights from Integrative Biophysical Modeling and Network Analysis of Conformational Landscapes.J Mol Biol. 2022 Sep 15;434(17):167506. doi: 10.1016/j.jmb.2022.167506. Epub 2022 Feb 21. J Mol Biol. 2022. PMID: 35202628
-
Extended conformational states dominate the Hsp90 chaperone dynamics.J Biol Chem. 2022 Jul;298(7):102101. doi: 10.1016/j.jbc.2022.102101. Epub 2022 Jun 3. J Biol Chem. 2022. PMID: 35667441 Free PMC article.
-
Allosteric Regulation Points Control the Conformational Dynamics of the Molecular Chaperone Hsp90.J Mol Biol. 2016 Nov 6;428(22):4559-4571. doi: 10.1016/j.jmb.2016.09.014. Epub 2016 Sep 20. J Mol Biol. 2016. PMID: 27663270
-
The Hsp90 chaperone machinery: conformational dynamics and regulation by co-chaperones.Biochim Biophys Acta. 2012 Mar;1823(3):624-35. doi: 10.1016/j.bbamcr.2011.09.003. Epub 2011 Sep 16. Biochim Biophys Acta. 2012. PMID: 21951723 Review.
-
Heat-shock protein 90, a chaperone for folding and regulation.Cell Mol Life Sci. 2002 Oct;59(10):1640-8. doi: 10.1007/pl00012491. Cell Mol Life Sci. 2002. PMID: 12475174 Free PMC article. Review.
Cited by
-
How specific ion effects influence the mechanical behaviors of amide macromolecules? A cross-scale study.RSC Adv. 2024 Aug 13;14(35):25507-25515. doi: 10.1039/d4ra04360j. eCollection 2024 Aug 12. RSC Adv. 2024. PMID: 39139238 Free PMC article.
-
One-pot dual protein labeling for simultaneous mechanical and fluorescent readouts in optical tweezers.Protein Sci. 2025 Apr;34(4):e70098. doi: 10.1002/pro.70098. Protein Sci. 2025. PMID: 40099877 Free PMC article.
-
A multiscale in situ time-resolved study of the nano- to millisecond structural dynamics during protein crystallization.J Appl Crystallogr. 2025 May 29;58(Pt 3):845-858. doi: 10.1107/S160057672500353X. eCollection 2025 Jun 1. J Appl Crystallogr. 2025. PMID: 40475925 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous