

This is a preprint.
Semi-synthetic nanobody-ligand conjugates exhibit tunable signaling properties and enhanced transcriptional outputs at neurokinin receptor-1
- PMID: 37986858
- PMCID: PMC10659424
- DOI: 10.1101/2023.10.08.561411
Semi-synthetic nanobody-ligand conjugates exhibit tunable signaling properties and enhanced transcriptional outputs at neurokinin receptor-1
Update in
-
Semi-synthetic nanobody-ligand conjugates exhibit tunable signaling properties and enhanced transcriptional outputs at neurokinin receptor-1.Protein Sci. 2024 Feb;33(2):e4866. doi: 10.1002/pro.4866. Protein Sci. 2024. PMID: 38088474 Free PMC article.
Abstract
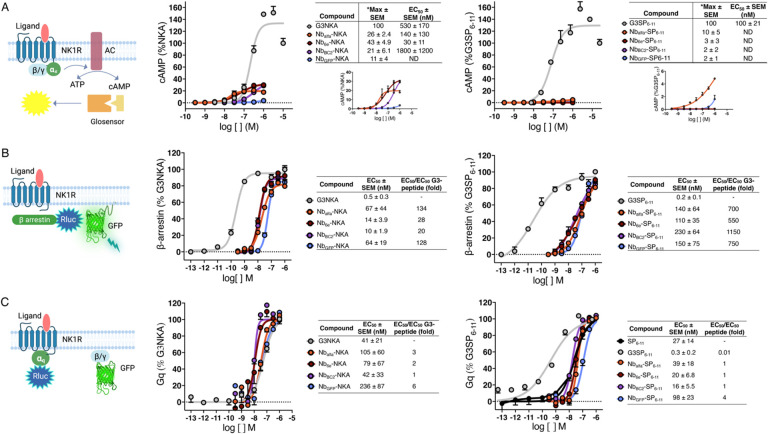
Antibodies have proven highly valuable for therapeutic development; however, they are typically poor candidates for applications that require activation of G protein-coupled receptors (GPCRs), the largest collection of targets for clinically approved drugs. Nanobodies (Nbs), the smallest antibody fragments retaining full antigen-binding capacity, have emerged as promising tools for pharmacologic applications, including GPCR modulation. Past work has shown that conjugation of Nbs with ligands can provide GPCR agonists that exhibit improved activity and selectivity compared to their parent ligands. The neurokinin-1 receptor (NK1R), a GPCR targeted for the treatment of pain, is activated by peptide agonists such as Substance P (SP) and neurokinin A (NKA), which induce signaling through multiple pathways (Gs, Gq and β-arrestin). In this study, we investigated whether conjugating NK1R ligands with Nbs that bind to a separate location on the receptor would provide chimeric compounds with distinctive signaling properties. We employed sortase A-mediated ligation to generate several conjugates consisting of Nbs linked to NK1R ligands. Many of these conjugates exhibited divergent and unexpected signaling properties and transcriptional outputs. For example, some Nb-NKA conjugates showed enhanced receptor binding capacity, high potency partial agonism, prolonged cAMP production, and an increase in transcriptional output associated with Gs signaling; whereas other conjugates were virtually inactive. Nanobody conjugation caused only minor alterations in ligand-induced upstream Gq signaling with unexpected enhancements in transcriptional (downstream) responses. Our findings underscore the potential of nanobody conjugation for providing compounds with advantageous properties such as biased agonism, prolonged duration of action, and enhanced transcriptional responses. These compounds hold promise not only for facilitating fundamental research on GPCR signal transduction mechanisms but also for the development of more potent and enduring therapeutics.
Conflict of interest statement
Declaration of interests: The authors declare no competing interests.
Figures




References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous