

Galectin-3 aggravates microglial activation and tau transmission in tauopathy
- PMID: 37988169
- PMCID: PMC10786694
- DOI: 10.1172/JCI165523
Galectin-3 aggravates microglial activation and tau transmission in tauopathy
Abstract
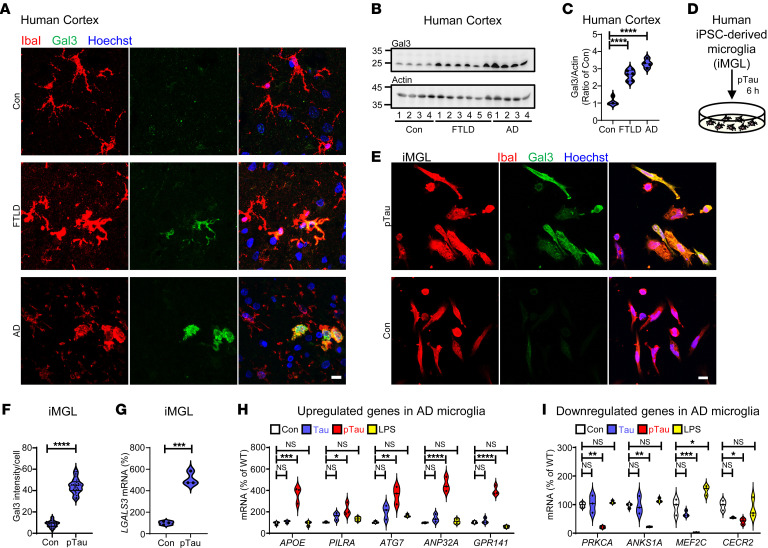
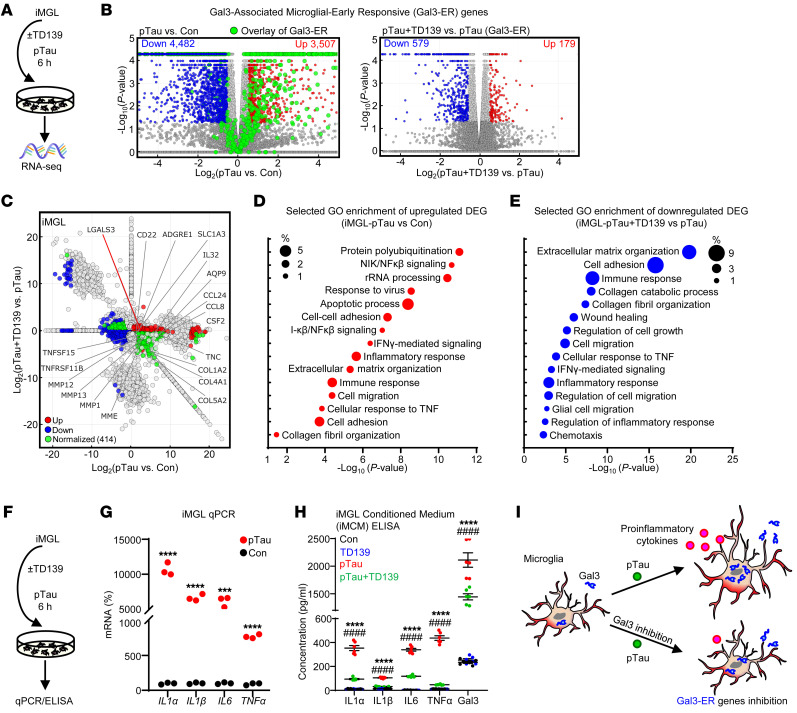
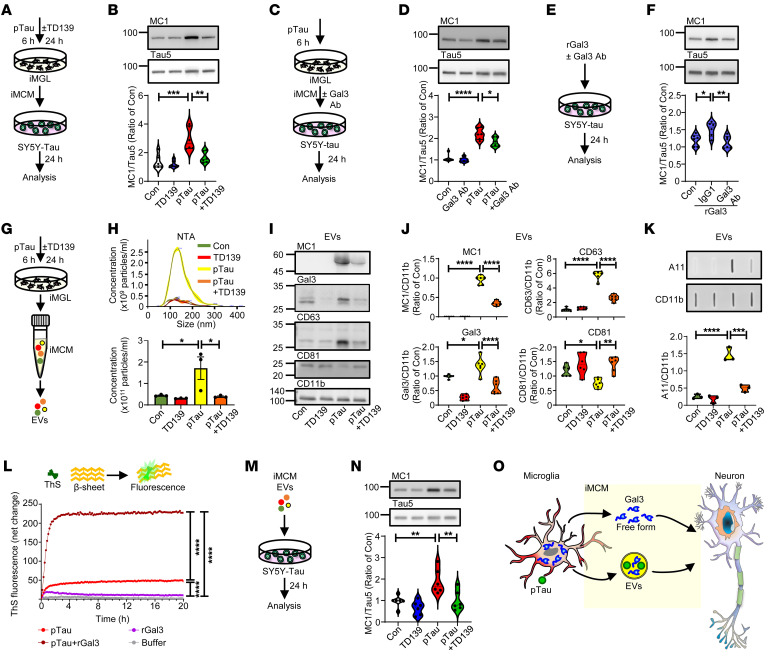
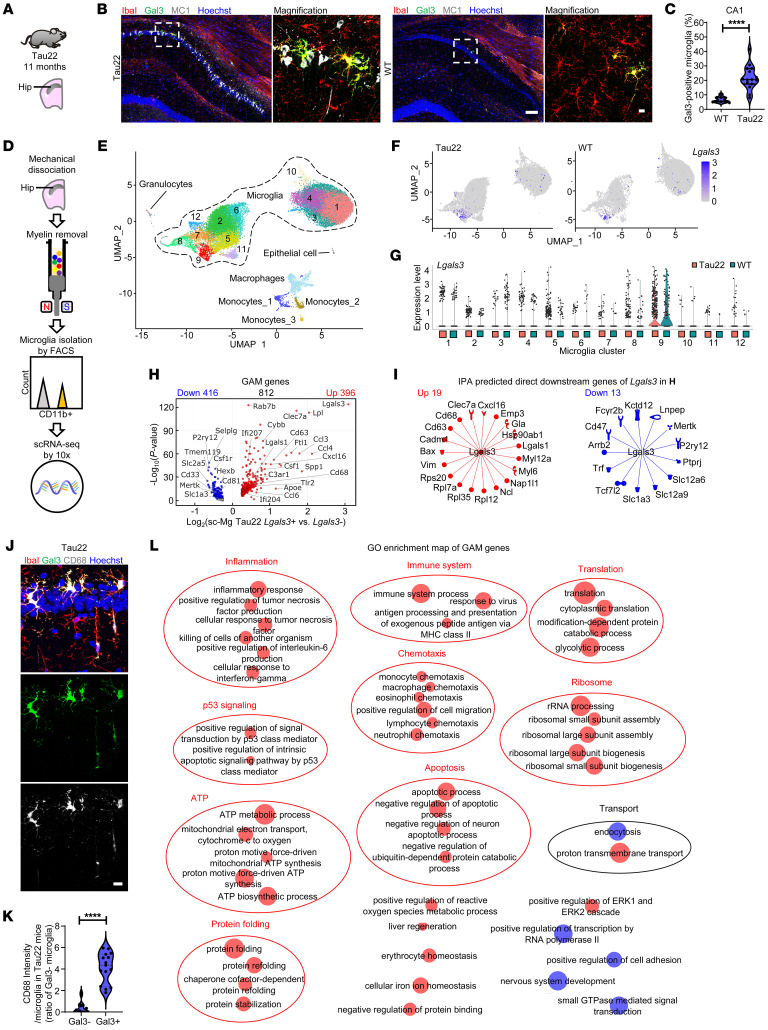
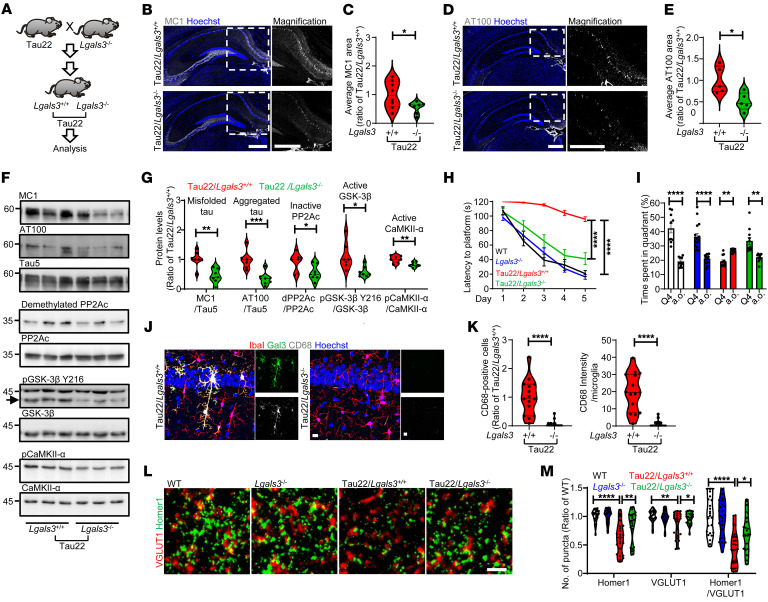
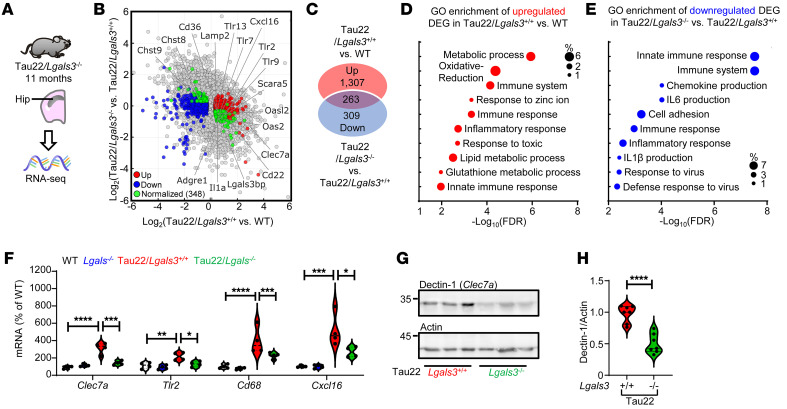
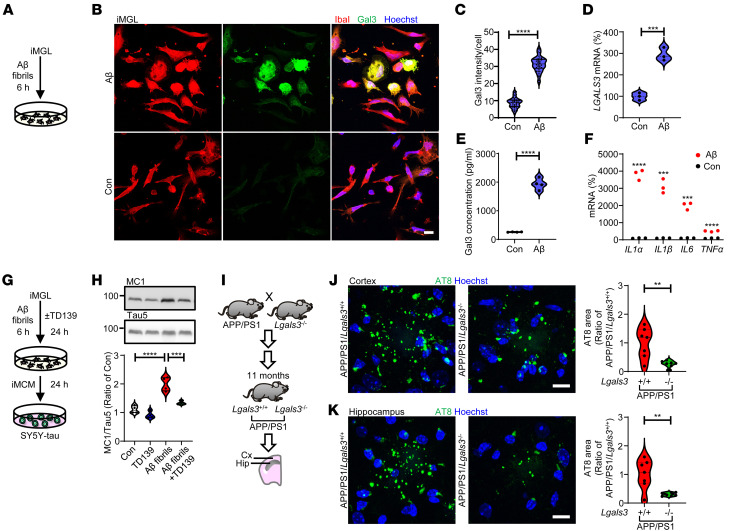
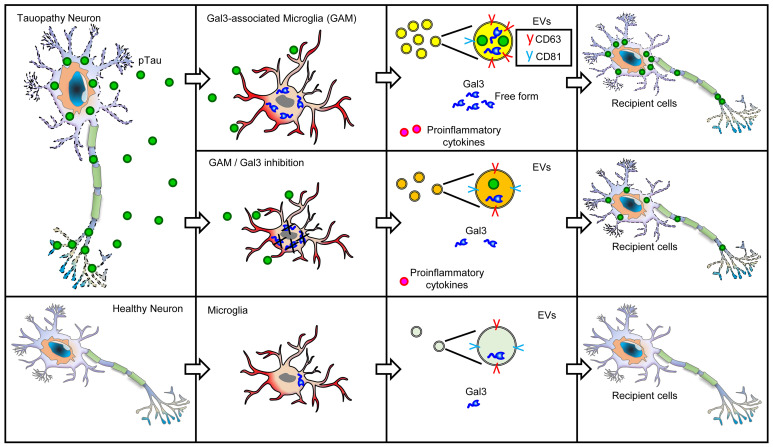
Alzheimer's disease is characterized by the accumulation of amyloid-β plaques, aggregation of hyperphosphorylated tau (pTau), and microglia activation. Galectin-3 (Gal3) is a β-galactoside-binding protein that has been implicated in amyloid pathology. Its role in tauopathy remains enigmatic. Here, we showed that Gal3 was upregulated in the microglia of humans and mice with tauopathy. pTau triggered the release of Gal3 from human induced pluripotent stem cell-derived microglia in both its free and extracellular vesicular-associated (EV-associated) forms. Both forms of Gal3 increased the accumulation of pathogenic tau in recipient cells. Binding of Gal3 to pTau greatly enhanced tau fibrillation. Besides Gal3, pTau was sorted into EVs for transmission. Moreover, pTau markedly enhanced the number of EVs released by iMGL in a Gal3-dependent manner, suggesting a role of Gal3 in biogenesis of EVs. Single-cell RNA-Seq analysis of the hippocampus of a mouse model of tauopathy (THY-Tau22) revealed a group of pathogenic tau-evoked, Gal3-associated microglia with altered cellular machineries implicated in neurodegeneration, including enhanced immune and inflammatory responses. Genetic removal of Gal3 in THY-Tau22 mice suppressed microglia activation, reduced the level of pTau and synaptic loss in neurons, and rescued memory impairment. Collectively, Gal3 is a potential therapeutic target for tauopathy.
Keywords: Alzheimer disease; Neurodegeneration; Neuroscience; iPS cells.
Conflict of interest statement
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
