

Solanum lycopersicum heme-binding protein 2 as a potent antimicrobial weapon against plant pathogens
- PMID: 37990046
- PMCID: PMC10663603
- DOI: 10.1038/s41598-023-47236-z
Solanum lycopersicum heme-binding protein 2 as a potent antimicrobial weapon against plant pathogens
Abstract
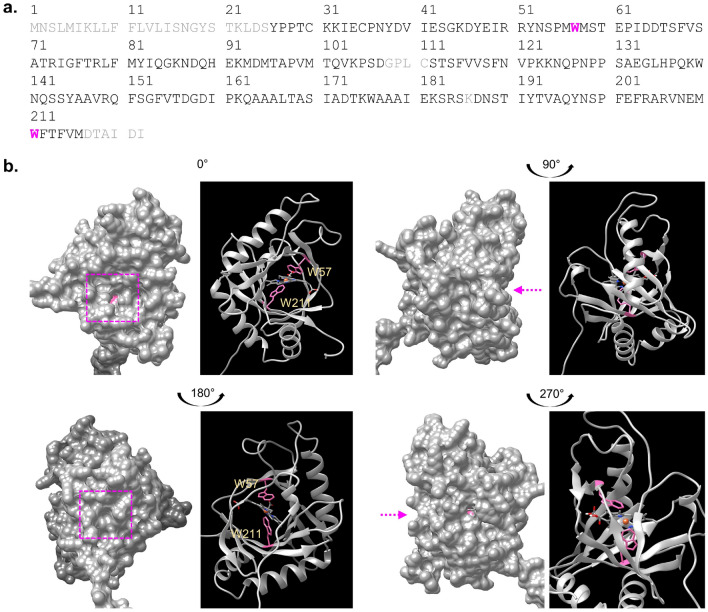
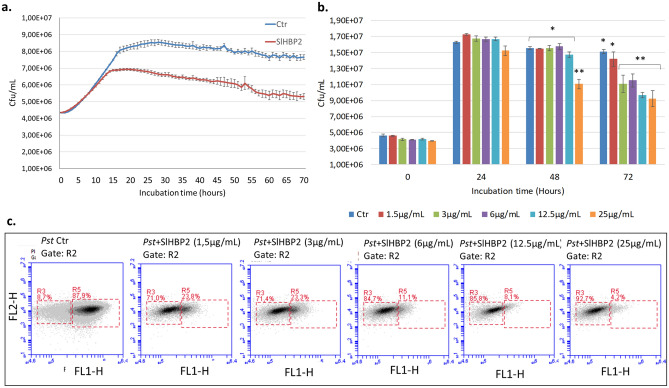
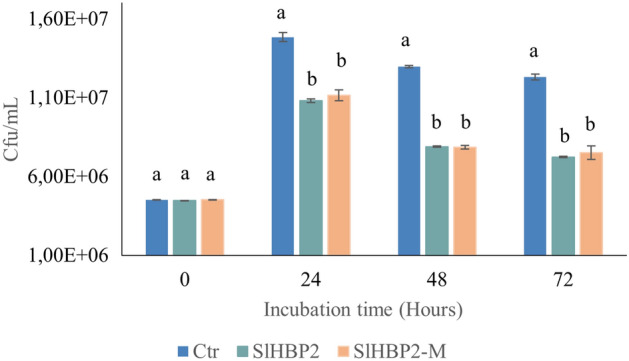
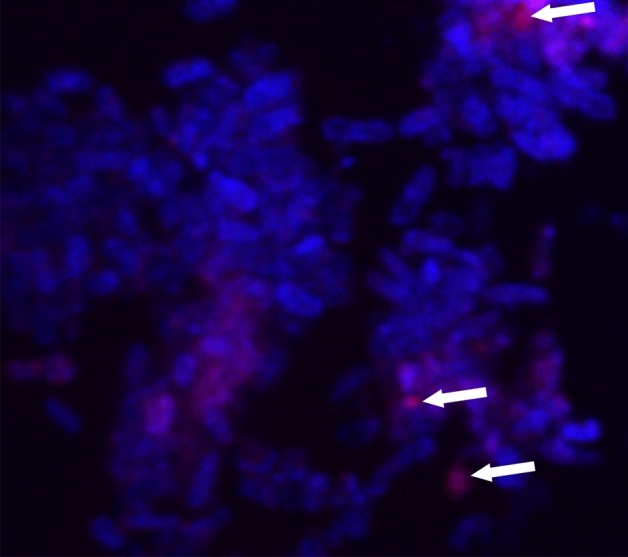
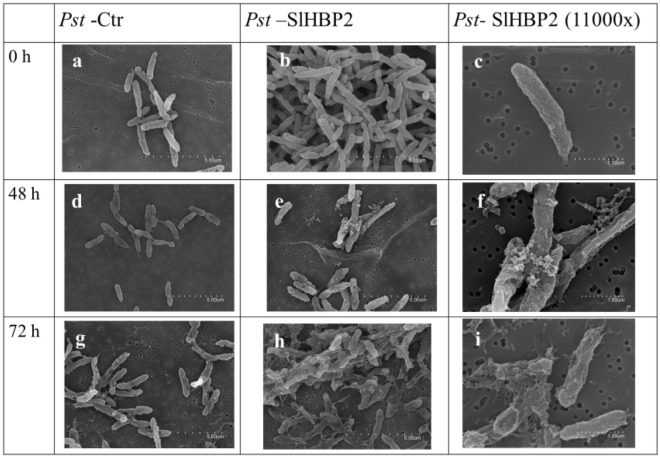
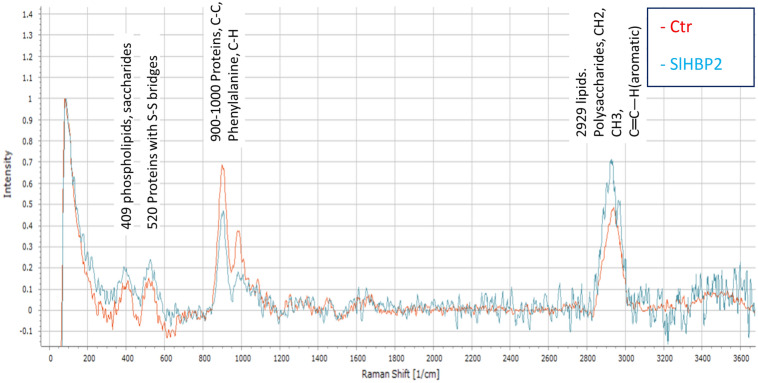
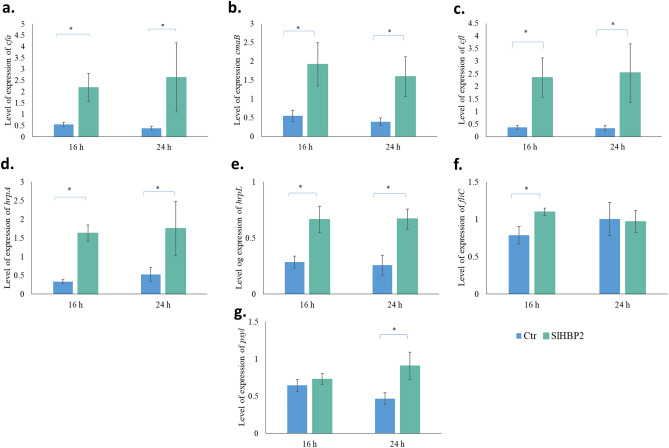
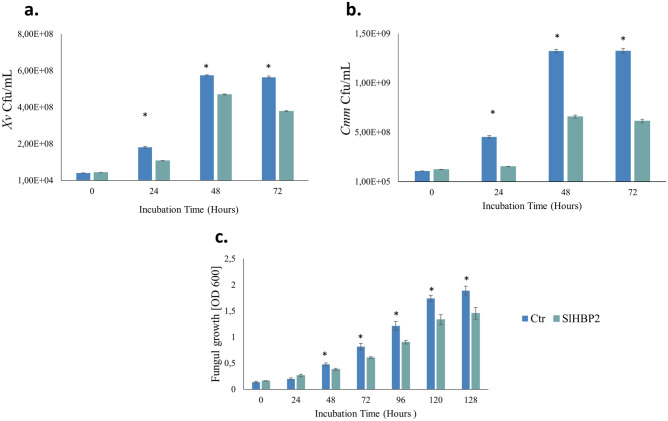
The rise in antibiotic-resistant bacteria caused by the excessive use of antibiotics has led to the urgent exploration of alternative antimicrobial solutions. Among these alternatives, antimicrobial proteins, and peptides (Apps) have garnered attention due to their wide-ranging antimicrobial effects. This study focuses on evaluating the antimicrobial properties of Solanum lycopersicum heme-binding protein 2 (SlHBP2), an apoplastic protein extracted from tomato plants treated with 1-Methyl tryptophan (1-MT), against Pseudomonas syringae pv. tomato DC3000 (Pst). Computational studies indicate that SlHBP2 is annotated as a SOUL heme-binding family protein. Remarkably, recombinant SlHBP2 demonstrated significant efficacy in inhibiting the growth of Pst within a concentration range of 3-25 μg/mL. Moreover, SlHBP2 exhibited potent antimicrobial effects against other microorganisms, including Xanthomonas vesicatoria (Xv), Clavibacter michiganensis subsp. michiganensis (Cmm), and Botrytis cinerea. To understand the mechanism of action employed by SlHBP2 against Pst, various techniques such as microscopy and fluorescence assays were employed. The results revealed that SlHBP2 disrupts the bacterial cell wall and causes leakage of intracellular contents. To summarize, the findings suggest that SlHBP2 has significant antimicrobial properties, making it a potential antimicrobial agent against a wide range of pathogens. Although further studies are warranted to explore the full potential of SlHBP2 and its suitability in various applications.
© 2023. The Author(s).
Conflict of interest statement
The authors Loredana Scalschi; Begonya Vicedo; Eugenio Llorens; Atefeh Farvardin; Gemma Camañes; and Eva Falomir, are listed as inventors in the patent (ES 2 879 643 B2) entitled: Antimicrobial proteins and peptides with stress-resistance-inducing activity. Authors Luisa Liu-Xu, Lorena Sanchez-Gimenez, Aloysius Wong, Elena G. Biosca and Jose M. Pedra, declare no competing interests.
Figures
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
