

Circadian clock factors regulate the first condensation reaction of fatty acid synthesis in Arabidopsis
- PMID: 37995186
- PMCID: PMC10842715
- DOI: 10.1016/j.celrep.2023.113483
Circadian clock factors regulate the first condensation reaction of fatty acid synthesis in Arabidopsis
Abstract
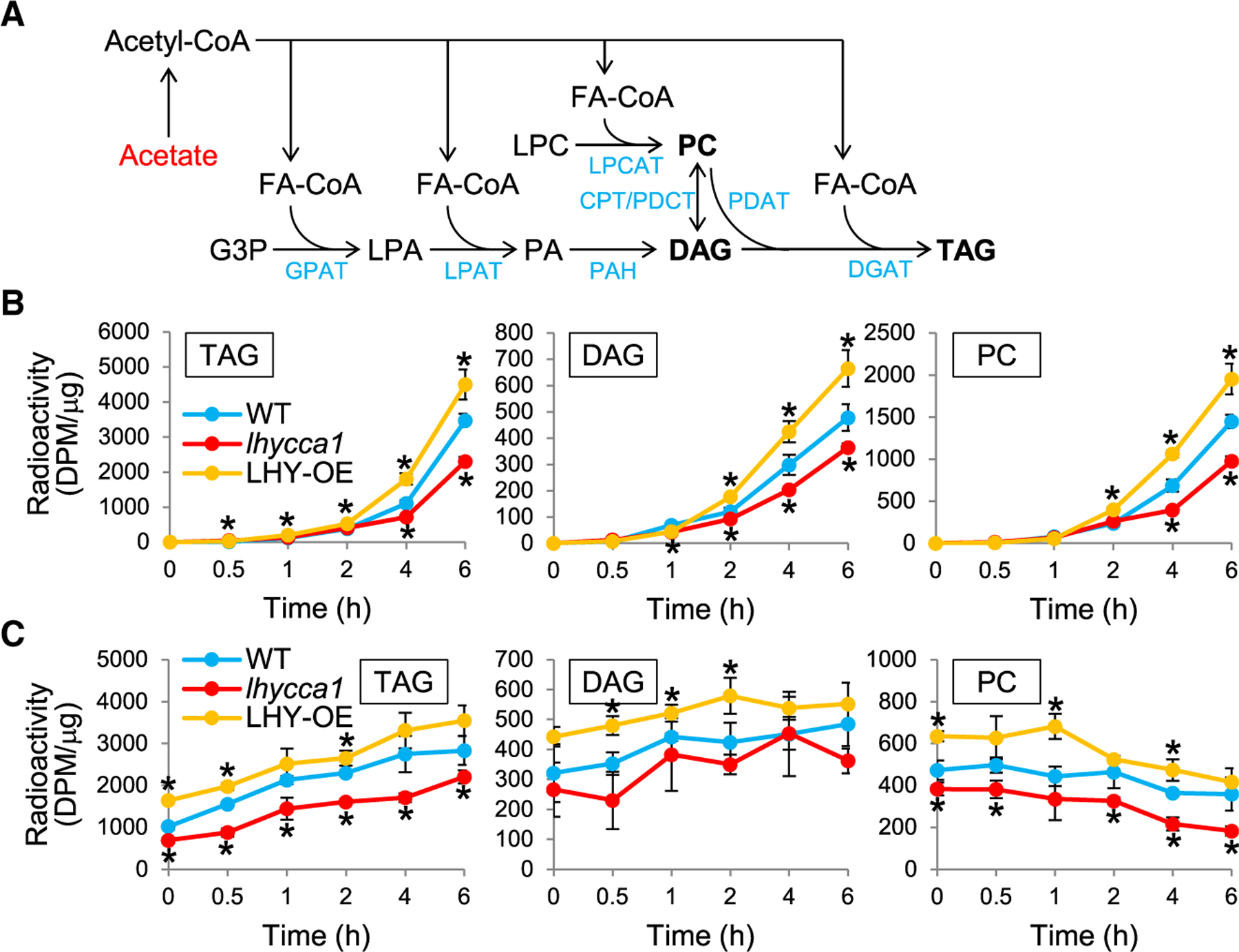
The circadian clock regulates temporal metabolic activities, but how it affects lipid metabolism is poorly understood. Here, we show that the central clock regulators LATE ELONGATED HYPOCOTYL (LHY) and CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) regulate the initial step of fatty acid (FA) biosynthesis in Arabidopsis. Triacylglycerol (TAG) accumulation in seeds was increased in LHY-overexpressing (LHY-OE) and decreased in lhycca1 plants. Metabolic tracking of lipids in developing seeds indicated that LHY enhanced FA synthesis. Transcript analysis revealed that the expression of genes involved in FA synthesis, including the one encoding β-ketoacyl-ACP synthase III (KASIII), was oppositely changed in developing seeds of LHY/CCA1-OEs and lhycca1. Chromatin immunoprecipitation, electrophoretic mobility shift, and transactivation assays indicated that LHY bound and activated the promoter of KASIII. Furthermore, phosphatidic acid, a metabolic precursor to TAG, inhibited LHY binding to KASIII promoter elements. Our data show a regulatory mechanism for plant lipid biosynthesis by the molecular clock.
Keywords: Arabidopsis; CP: Plants; Clock regulation of lipid metabolism; LHY/CCA1; Lipid-clock interconnection; b-ketoacyl-ACP synthase III (KASIII); circadian clock; fatty acid synthesis; lipid signaling; phosphatidic acid; seed oil; transcriptional regulation.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





Similar articles
-
LATE ELONGATED HYPOCOTYL regulates photoperiodic flowering via the circadian clock in Arabidopsis.BMC Plant Biol. 2016 May 20;16(1):114. doi: 10.1186/s12870-016-0810-8. BMC Plant Biol. 2016. PMID: 27207270 Free PMC article.
-
CIRCADIAN CLOCK ASSOCIATED1 and LATE ELONGATED HYPOCOTYL function synergistically in the circadian clock of Arabidopsis.Plant Physiol. 2009 Jun;150(2):834-43. doi: 10.1104/pp.108.133272. Epub 2009 Feb 13. Plant Physiol. 2009. PMID: 19218364 Free PMC article.
-
The central circadian clock proteins CCA1 and LHY regulate iron homeostasis in Arabidopsis.J Integr Plant Biol. 2019 Feb;61(2):168-181. doi: 10.1111/jipb.12696. Epub 2018 Aug 31. J Integr Plant Biol. 2019. PMID: 29989313
-
MYB transcription factors in the Arabidopsis circadian clock.J Exp Bot. 2002 Jul;53(374):1551-7. doi: 10.1093/jxb/erf027. J Exp Bot. 2002. PMID: 12096093 Review.
-
The circadian system of Arabidopsis thaliana: forward and reverse genetic approaches.Chronobiol Int. 1999 Jan;16(1):1-16. doi: 10.3109/07420529908998708. Chronobiol Int. 1999. PMID: 10023572 Review.
Cited by
-
Thaumatin-like Gene TLP1b Confers to Seed Oil Content and Resistance to Sclerotinia sclerotiorum in Arabidopsis.Int J Mol Sci. 2025 Feb 24;26(5):1930. doi: 10.3390/ijms26051930. Int J Mol Sci. 2025. PMID: 40076556 Free PMC article.
-
Phosphatidic acid signaling in modulating plant reproduction and architecture.Plant Commun. 2025 Feb 10;6(2):101234. doi: 10.1016/j.xplc.2024.101234. Epub 2024 Dec 24. Plant Commun. 2025. PMID: 39722455 Free PMC article. Review.
-
Rhythmic lipid and gene expression responses to chilling in panicoid grasses.J Exp Bot. 2024 Sep 27;75(18):5790-5804. doi: 10.1093/jxb/erae247. J Exp Bot. 2024. PMID: 38808657 Free PMC article.
-
The circadian clock regulates receptor-mediated immune responses to an herbivore-associated molecular pattern.bioRxiv [Preprint]. 2024 Nov 8:2024.11.06.622352. doi: 10.1101/2024.11.06.622352. bioRxiv. 2024. PMID: 39574761 Free PMC article. Preprint.
-
Transcriptional activation and repression in the plant circadian clock: Revisiting core oscillator feedback loops and output pathways.Plant Commun. 2025 Aug 11;6(8):101415. doi: 10.1016/j.xplc.2025.101415. Epub 2025 Jun 10. Plant Commun. 2025. PMID: 40500965 Free PMC article. Review.
References
-
- Alabadí D, Oyama T, Yanovsky MJ, Harmon FG, Más P, and Kay SA (2001). Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293, 880–883. - PubMed
-
- Huang W, Pérez-García P, Pokhilko A, Millar AJ, Antoshechkin I, Riechmann JL, and Mas P (2012). Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator. Science 336, 75–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
