

Lineage-specific 3D genome organization is assembled at multiple scales by IKAROS
- PMID: 37995656
- PMCID: PMC10895928
- DOI: 10.1016/j.cell.2023.10.023
Lineage-specific 3D genome organization is assembled at multiple scales by IKAROS
Abstract
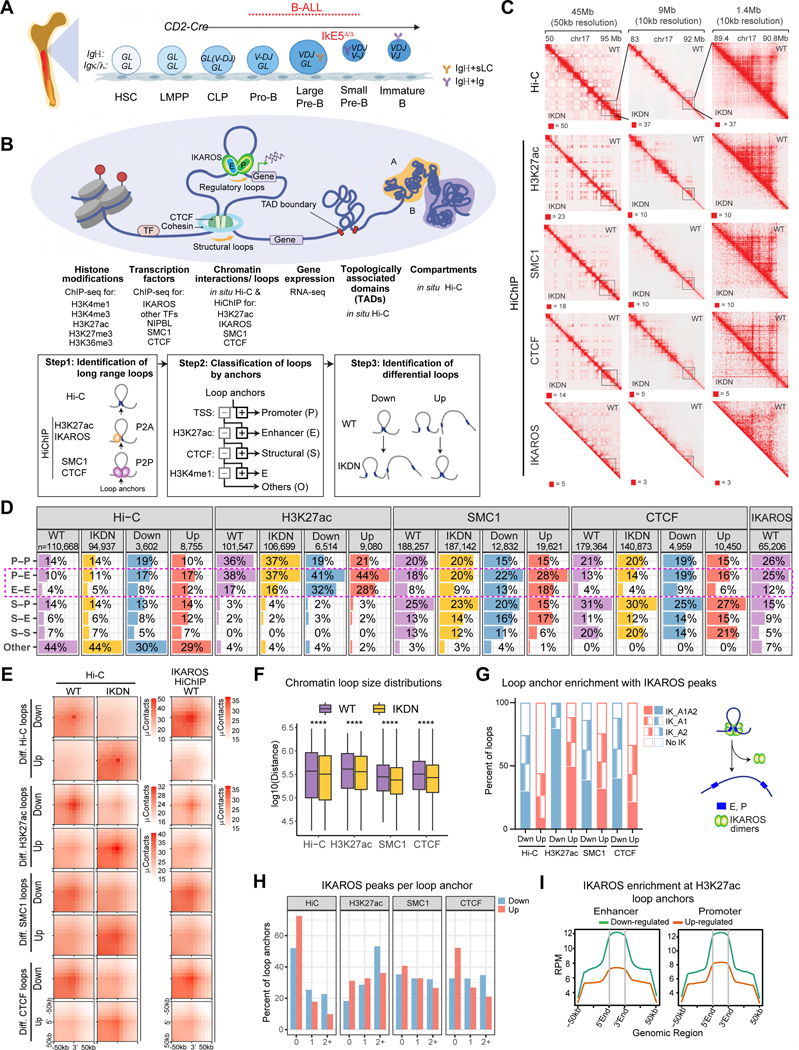
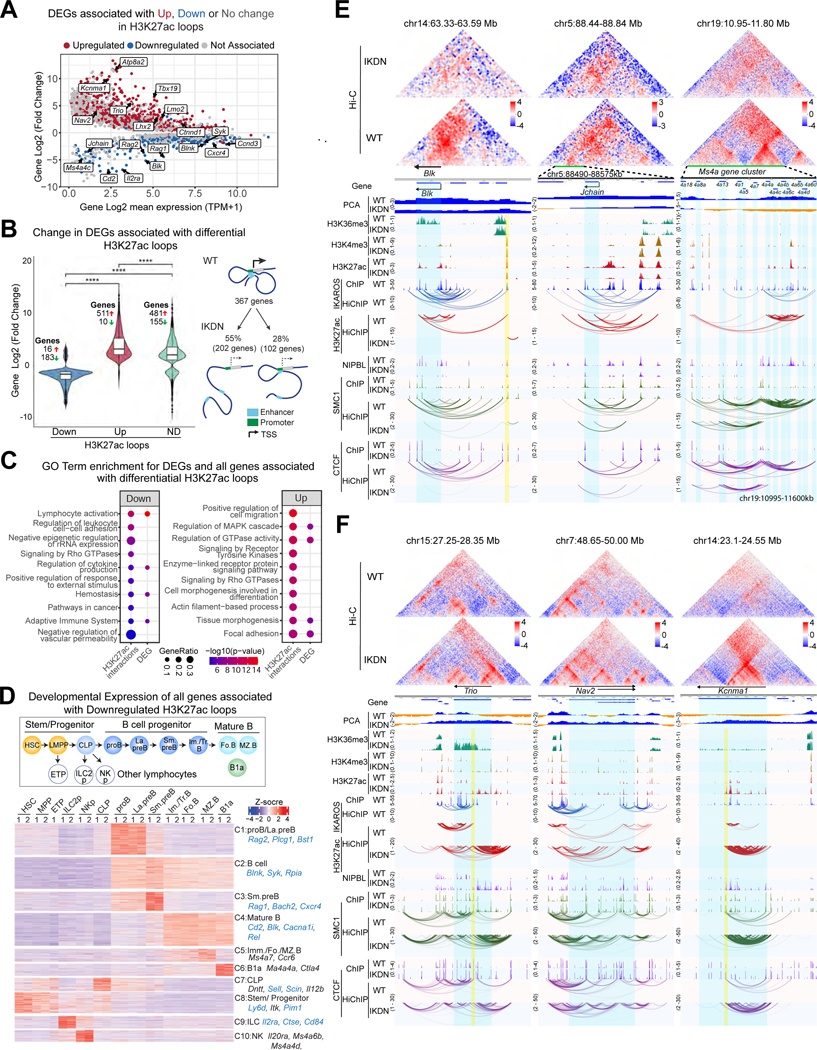
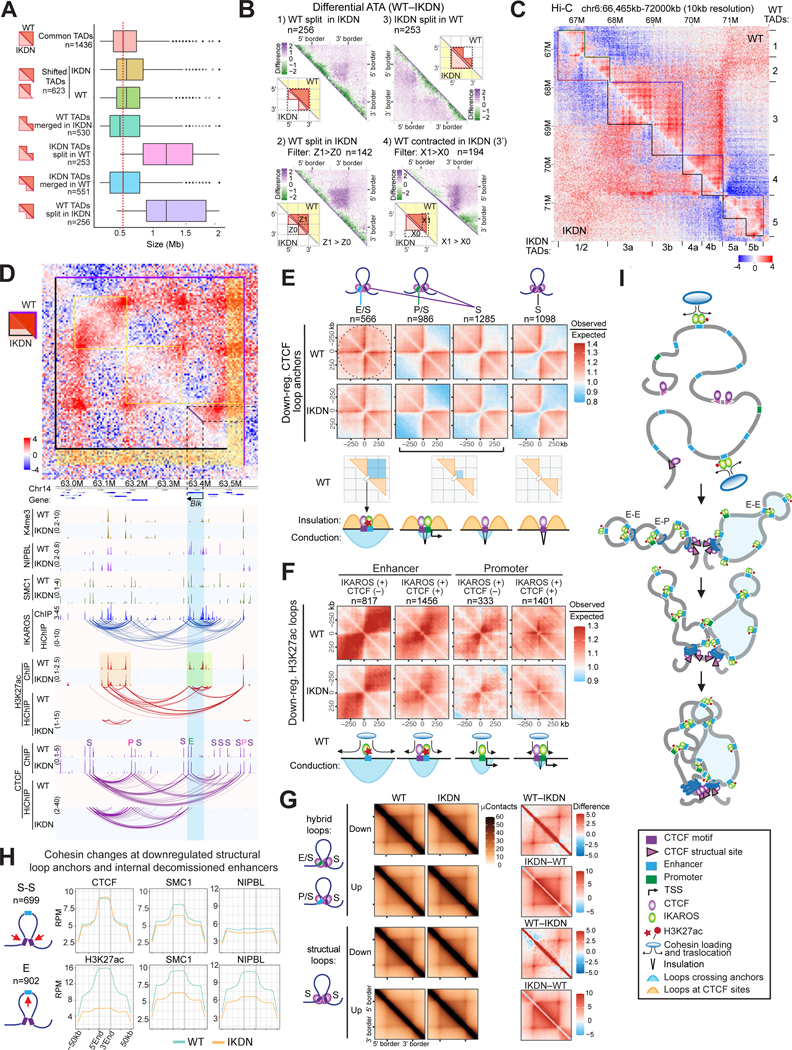
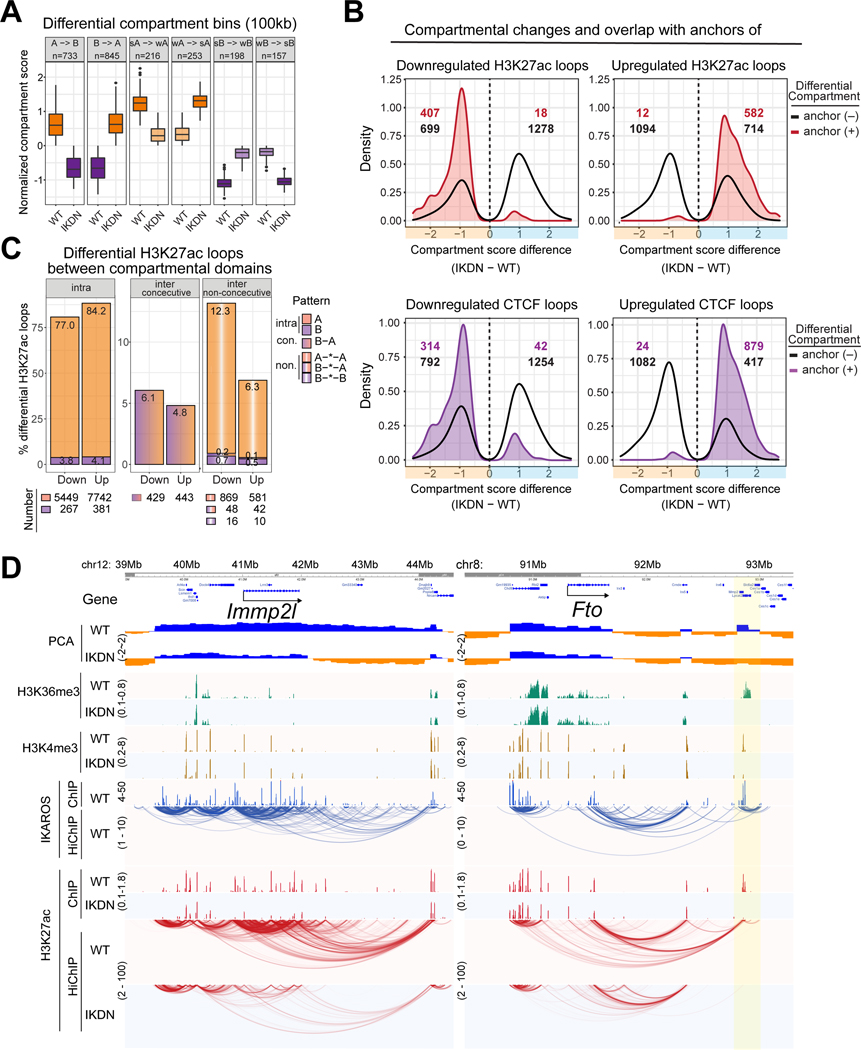
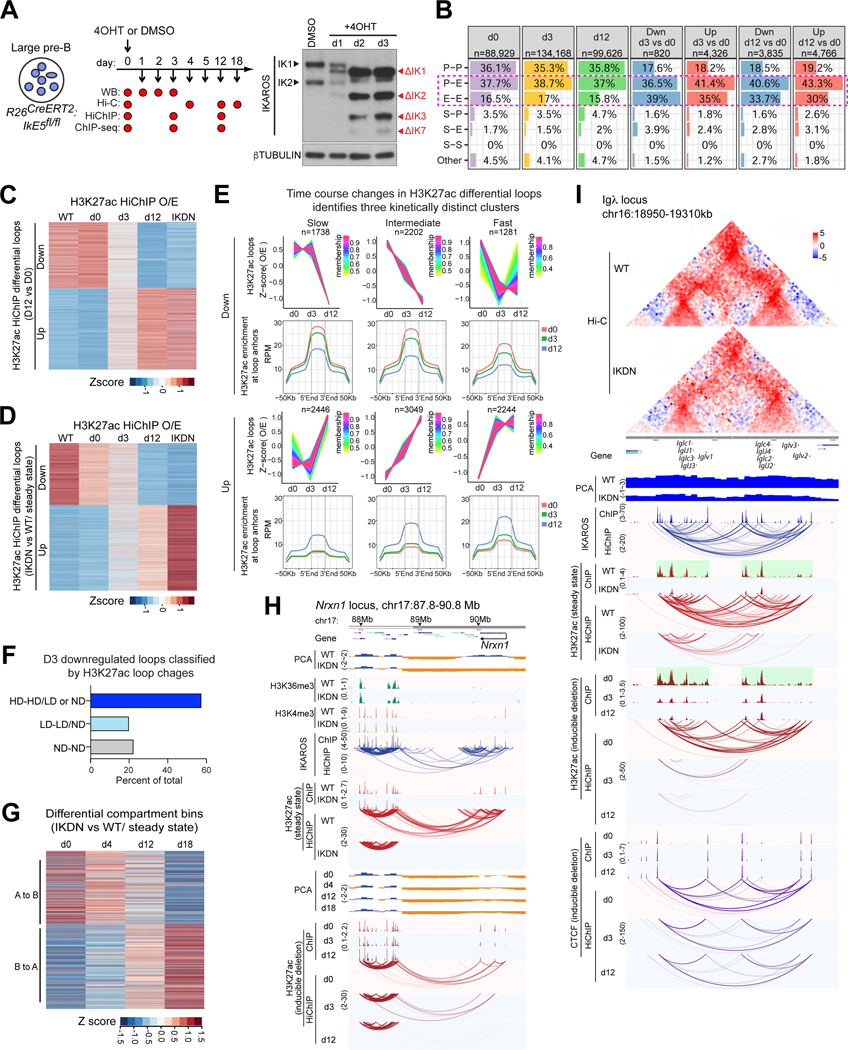
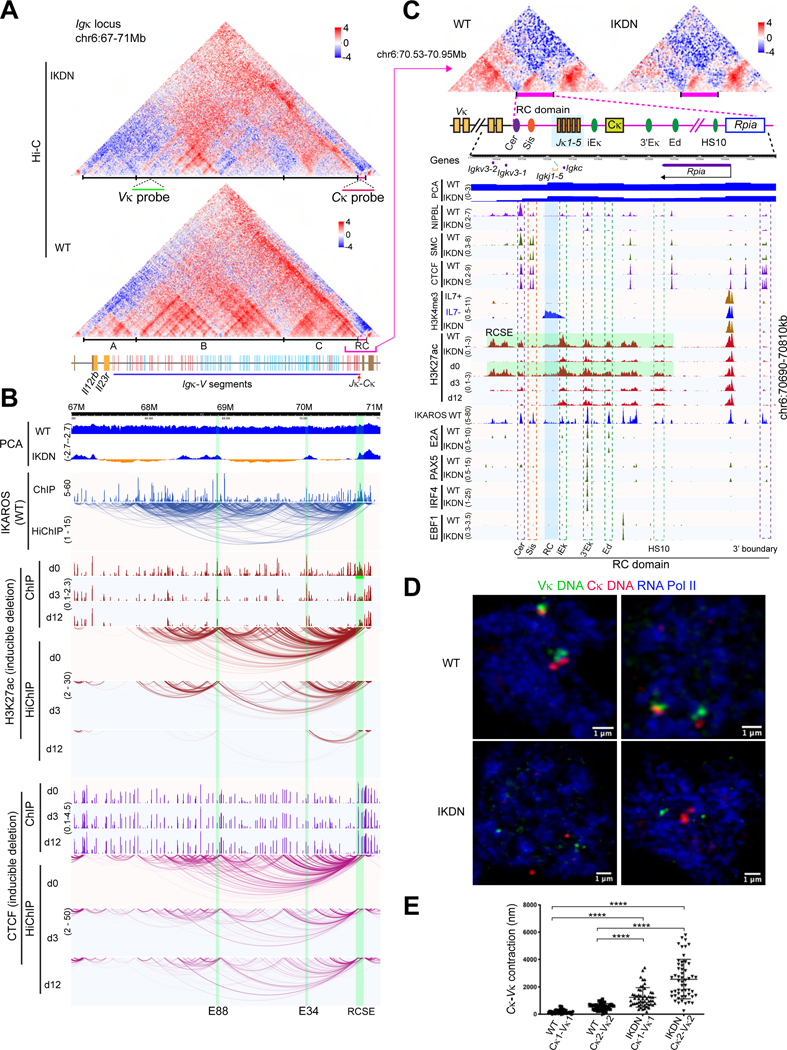
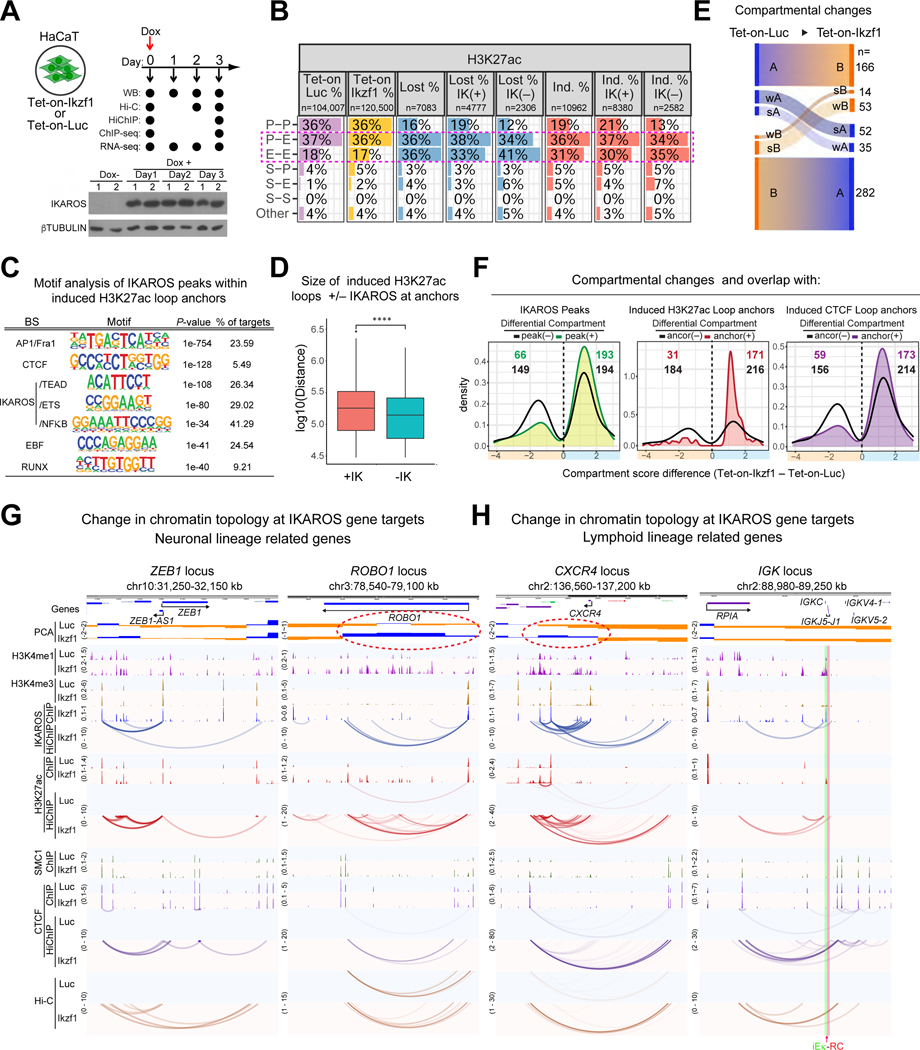
A generic level of chromatin organization generated by the interplay between cohesin and CTCF suffices to limit promiscuous interactions between regulatory elements, but a lineage-specific chromatin assembly that supersedes these constraints is required to configure the genome to guide gene expression changes that drive faithful lineage progression. Loss-of-function approaches in B cell precursors show that IKAROS assembles interactions across megabase distances in preparation for lymphoid development. Interactions emanating from IKAROS-bound enhancers override CTCF-imposed boundaries to assemble lineage-specific regulatory units built on a backbone of smaller invariant topological domains. Gain of function in epithelial cells confirms IKAROS' ability to reconfigure chromatin architecture at multiple scales. Although the compaction of the Igκ locus required for genome editing represents a function of IKAROS unique to lymphocytes, the more general function to preconfigure the genome to support lineage-specific gene expression and suppress activation of extra-lineage genes provides a paradigm for lineage restriction.
Keywords: 3D genome organization; Hi-C; HiChIP; IKAROS; Igκ locus contraction; enhancer loops; inter-compartmental loops; lineage-specific genome organization; lymphocyte development; superTADs.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
