

Perinatal foodborne titanium dioxide exposure-mediated dysbiosis predisposes mice to develop colitis through life
- PMID: 37996842
- PMCID: PMC10666382
- DOI: 10.1186/s12989-023-00555-5
Perinatal foodborne titanium dioxide exposure-mediated dysbiosis predisposes mice to develop colitis through life
Erratum in
-
Correction: Perinatal foodborne titanium dioxide exposure-mediated dysbiosis predisposes mice to develop colitis through life.Part Fibre Toxicol. 2024 Mar 6;21(1):11. doi: 10.1186/s12989-024-00570-0. Part Fibre Toxicol. 2024. PMID: 38448937 Free PMC article. No abstract available.
Abstract
Background: Perinatal exposure to titanium dioxide (TiO2), as a foodborne particle, may influence the intestinal barrier function and the susceptibility to develop inflammatory bowel disease (IBD) later in life. Here, we investigate the impact of perinatal foodborne TiO2 exposure on the intestinal mucosal function and the susceptibility to develop IBD-associated colitis. Pregnant and lactating mother mice were exposed to TiO2 until pups weaning and the gut microbiota and intestinal barrier function of their offspring was assessed at day 30 post-birth (weaning) and at adult age (50 days). Epigenetic marks was studied by DNA methylation profile measuring the level of 5-methyl-2'-deoxycytosine (5-Me-dC) in DNA from colic epithelial cells. The susceptibility to develop IBD has been monitored using dextran-sulfate sodium (DSS)-induced colitis model. Germ-free mice were used to define whether microbial transfer influence the mucosal homeostasis and subsequent exacerbation of DSS-induced colitis.
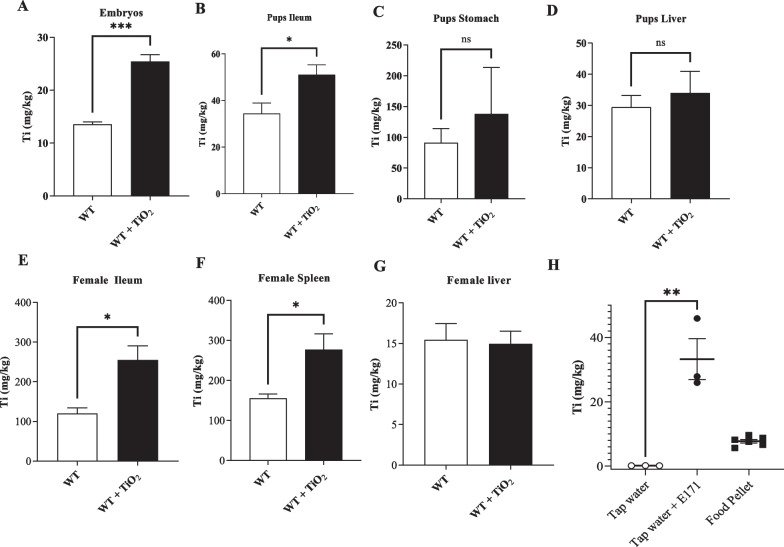
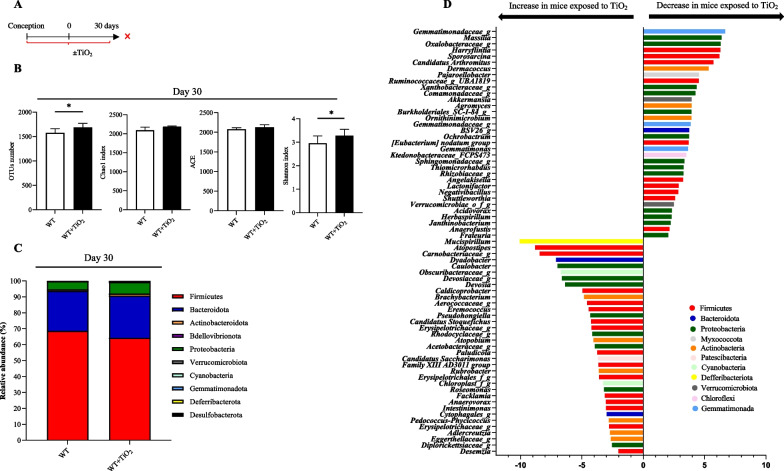
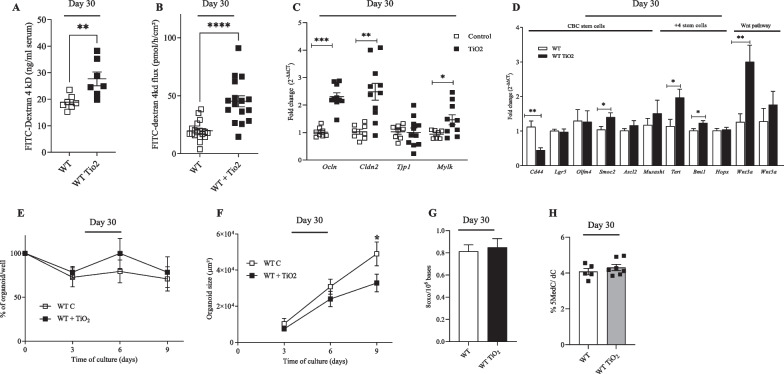
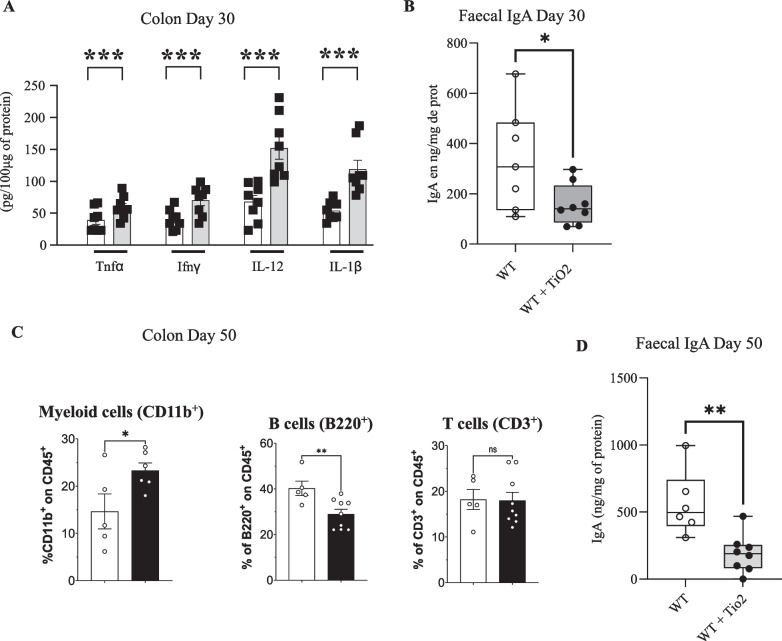
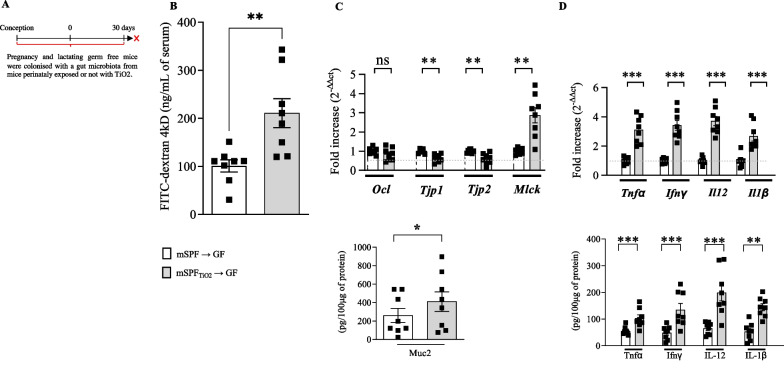
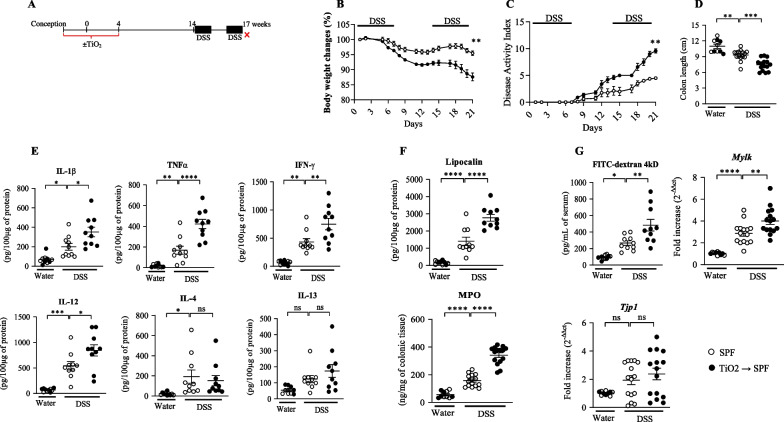
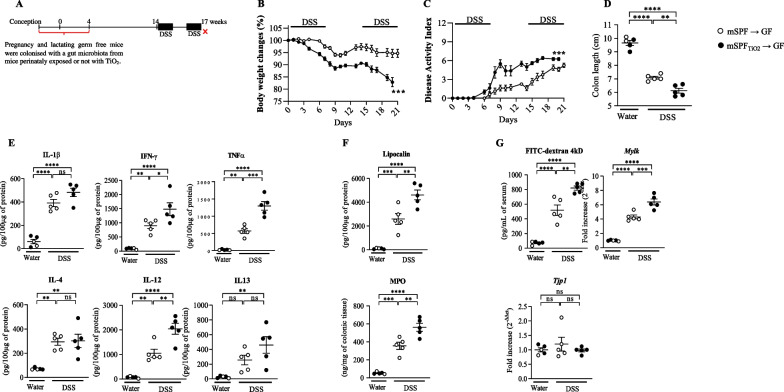
Results: In pregnant and lactating mice, foodborne TiO2 was able to translocate across the host barriers including gut, placenta and mammary gland to reach embryos and pups, respectively. This passage modified the chemical element composition of foetus, and spleen and liver of mothers and their offspring. We showed that perinatal exposure to TiO2 early in life alters the gut microbiota composition, increases the intestinal epithelial permeability and enhances the colonic cytokines and myosin light chain kinase expression. Moreover, perinatal exposure to TiO2 also modifies the abilities of intestinal stem cells to survive, grow and generate a functional epithelium. Maternal TiO2 exposure increases the susceptibility of offspring mice to develop severe DSS-induced colitis later in life. Finally, transfer of TiO2-induced microbiota dysbiosis to pregnant germ-free mice affects the homeostasis of the intestinal mucosal barrier early in life and confers an increased susceptibility to develop colitis in adult offspring.
Conclusions: Our findings indicate that foodborne TiO2 consumption during the perinatal period has negative long-lasting consequences on the development of the intestinal mucosal barrier toward higher colitis susceptibility. This demonstrates to which extent environmental factors influence the microbial-host interplay and impact the long-term mucosal homeostasis.
Keywords: Colitis; Foodborne TiO2; Intestinal barrier function; Intestinal stem cells; Microbiota; Perinatal period.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Prenatal Maternal Stress Exacerbates Experimental Colitis of Offspring in Adulthood.Front Immunol. 2021 Nov 3;12:700995. doi: 10.3389/fimmu.2021.700995. eCollection 2021. Front Immunol. 2021. PMID: 34804005 Free PMC article.
-
Maternal High Fat Diet Alters Gut Microbiota of Offspring and Exacerbates DSS-Induced Colitis in Adulthood.Front Immunol. 2018 Nov 13;9:2608. doi: 10.3389/fimmu.2018.02608. eCollection 2018. Front Immunol. 2018. PMID: 30483266 Free PMC article.
-
Enteric Delivery of Regenerating Family Member 3 alpha Alters the Intestinal Microbiota and Controls Inflammation in Mice With Colitis.Gastroenterology. 2018 Mar;154(4):1009-1023.e14. doi: 10.1053/j.gastro.2017.11.003. Epub 2017 Nov 11. Gastroenterology. 2018. PMID: 29133078
-
Perinatal exposure to foodborne inorganic nanoparticles: A role in the susceptibility to food allergy?Front Allergy. 2022 Dec 5;3:1067281. doi: 10.3389/falgy.2022.1067281. eCollection 2022. Front Allergy. 2022. PMID: 36545344 Free PMC article. Review.
-
The Intestinal Barrier-Shielding the Body from Nano- and Microparticles in Our Diet.Metabolites. 2022 Mar 2;12(3):223. doi: 10.3390/metabo12030223. Metabolites. 2022. PMID: 35323666 Free PMC article. Review.
Cited by
-
Correction: Perinatal foodborne titanium dioxide exposure-mediated dysbiosis predisposes mice to develop colitis through life.Part Fibre Toxicol. 2024 Mar 6;21(1):11. doi: 10.1186/s12989-024-00570-0. Part Fibre Toxicol. 2024. PMID: 38448937 Free PMC article. No abstract available.
-
Validating Enteroid-Derived Monolayers from Murine Gut Organoids for Toxicological Testing of Inorganic Particles: Proof-of-Concept with Food-Grade Titanium Dioxide.Int J Mol Sci. 2024 Feb 23;25(5):2635. doi: 10.3390/ijms25052635. Int J Mol Sci. 2024. PMID: 38473881 Free PMC article.
-
Genome-wide DNA methylation sequencing reveals the involvement of ferroptosis in hepatotoxicity induced by dietary exposure to food-grade titanium dioxide.Part Fibre Toxicol. 2024 Sep 18;21(1):37. doi: 10.1186/s12989-024-00598-2. Part Fibre Toxicol. 2024. PMID: 39294687 Free PMC article.
