

Acute expression of human APOBEC3B in mice results in RNA editing and lethality
- PMID: 38001542
- PMCID: PMC10668425
- DOI: 10.1186/s13059-023-03115-4
Acute expression of human APOBEC3B in mice results in RNA editing and lethality
Abstract
Background: RNA editing has been described as promoting genetic heterogeneity, leading to the development of multiple disorders, including cancer. The cytosine deaminase APOBEC3B is implicated in tumor evolution through DNA mutation, but whether it also functions as an RNA editing enzyme has not been studied.
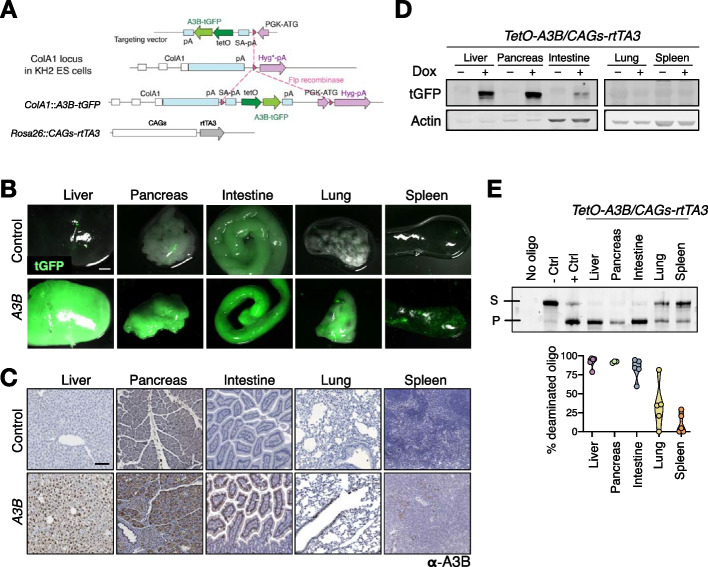
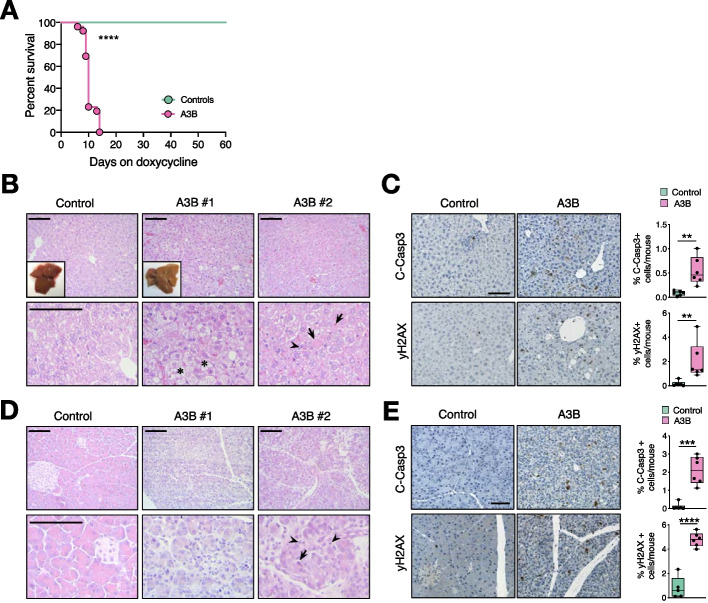
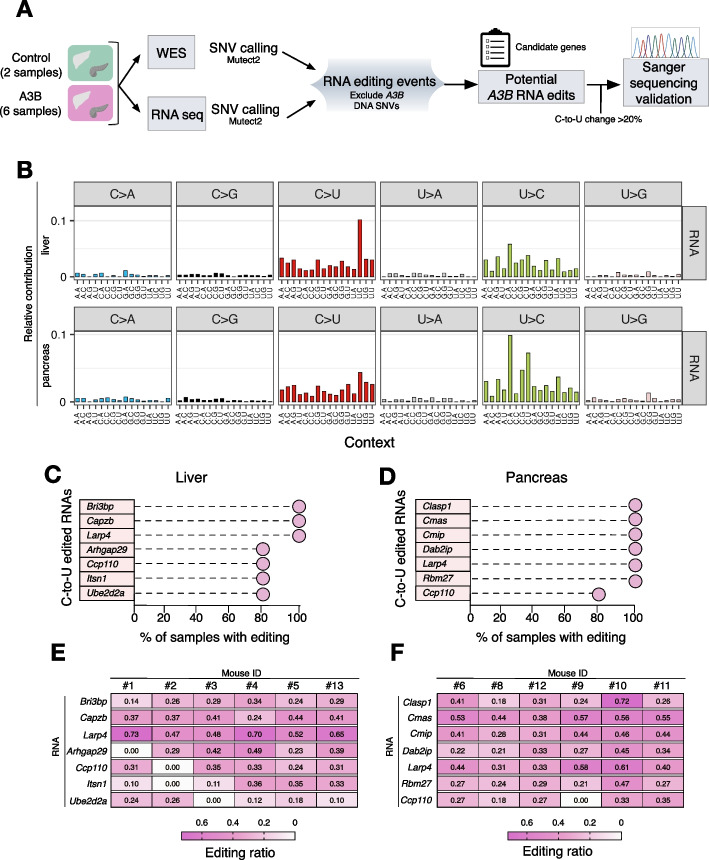
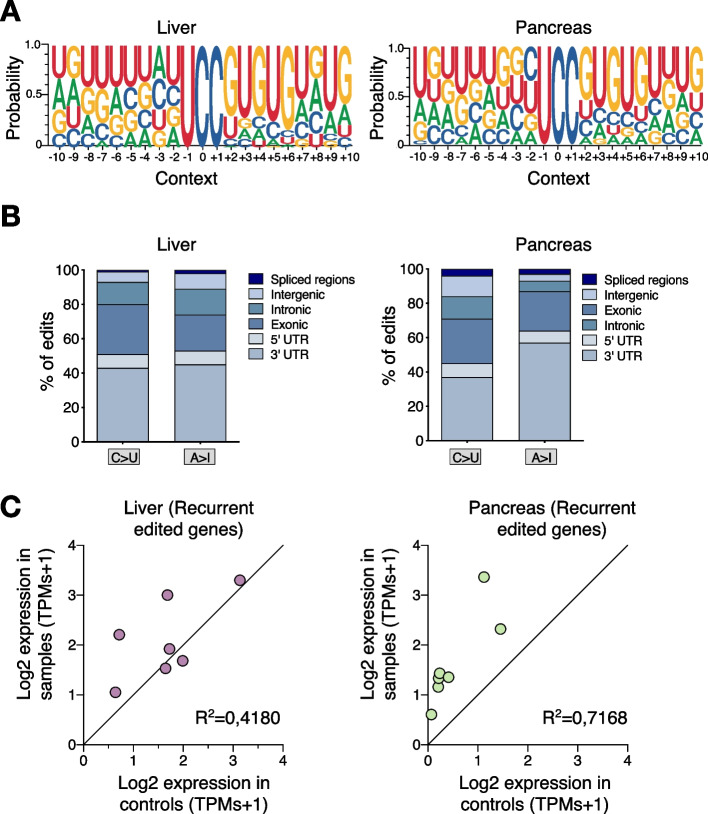
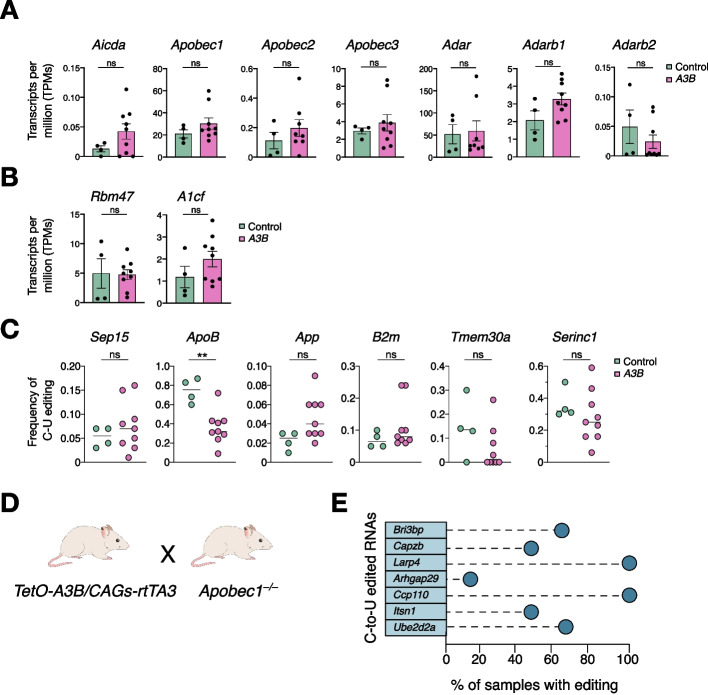
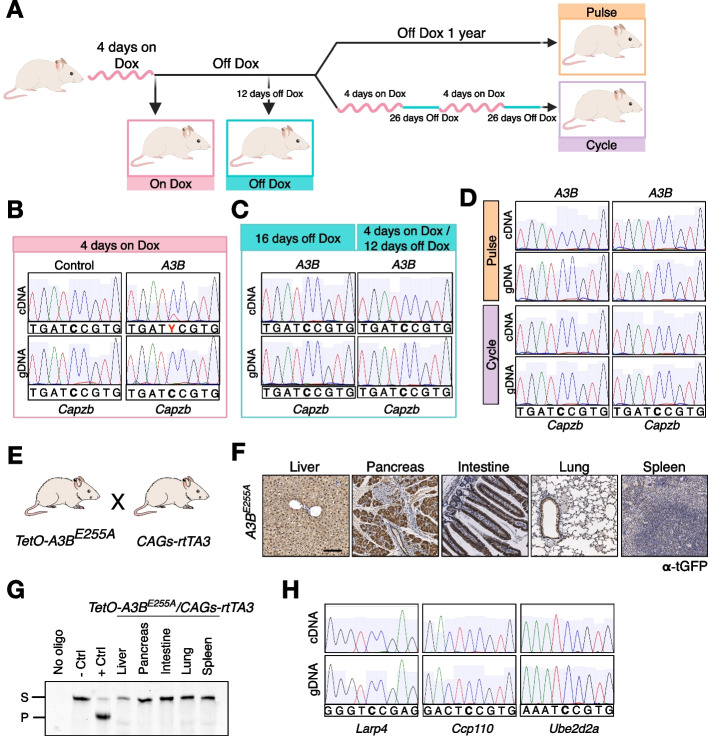
Results: Here, we engineer a novel doxycycline-inducible mouse model of human APOBEC3B-overexpression to understand the impact of this enzyme in tissue homeostasis and address a potential role in C-to-U RNA editing. Elevated and sustained levels of APOBEC3B lead to rapid alteration of cellular fitness, major organ dysfunction, and ultimately lethality in mice. Importantly, RNA-sequencing of mouse tissues expressing high levels of APOBEC3B identifies frequent UCC-to-UUC RNA editing events that are not evident in the corresponding genomic DNA.
Conclusions: This work identifies, for the first time, a new deaminase-dependent function for APOBEC3B in RNA editing and presents a preclinical tool to help understand the emerging role of APOBEC3B as a driver of carcinogenesis.
Keywords: APOBEC3B; DNA damage; Mouse models; Mutations; RNA editing.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
The DNA cytosine deaminase APOBEC3B promotes tamoxifen resistance in ER-positive breast cancer.Sci Adv. 2016 Oct 7;2(10):e1601737. doi: 10.1126/sciadv.1601737. eCollection 2016 Oct. Sci Adv. 2016. PMID: 27730215 Free PMC article.
-
Ancestral APOBEC3B Nuclear Localization Is Maintained in Humans and Apes and Altered in Most Other Old World Primate Species.mSphere. 2022 Dec 21;7(6):e0045122. doi: 10.1128/msphere.00451-22. Epub 2022 Nov 14. mSphere. 2022. PMID: 36374108 Free PMC article.
-
APOBEC3B: pathological consequences of an innate immune DNA mutator.Biomed J. 2015 Mar-Apr;38(2):102-10. doi: 10.4103/2319-4170.148904. Biomed J. 2015. PMID: 25566802 Review.
-
APOBEC3B edits HBV DNA and inhibits HBV replication during reverse transcription.Antiviral Res. 2018 Jan;149:16-25. doi: 10.1016/j.antiviral.2017.11.006. Epub 2017 Nov 10. Antiviral Res. 2018. PMID: 29129707
-
Roles of APOBEC3A and APOBEC3B in Human Papillomavirus Infection and Disease Progression.Viruses. 2017 Aug 21;9(8):233. doi: 10.3390/v9080233. Viruses. 2017. PMID: 28825669 Free PMC article. Review.
Cited by
-
Computation strategies and clinical applications in neoantigen discovery towards precision cancer immunotherapy.Biomark Res. 2025 Jul 9;13(1):96. doi: 10.1186/s40364-025-00808-9. Biomark Res. 2025. PMID: 40629481 Free PMC article. Review.
-
ATR safeguards replication forks against APOBEC3B-induced toxic PARP1 trapping.bioRxiv [Preprint]. 2024 Nov 15:2024.11.14.623607. doi: 10.1101/2024.11.14.623607. bioRxiv. 2024. Update in: Sci Adv. 2025 Apr 18;11(16):eadu0437. doi: 10.1126/sciadv.adu0437. PMID: 39605722 Free PMC article. Updated. Preprint.
-
Mesoscale DNA features impact APOBEC3A and APOBEC3B deaminase activity and shape tumor mutational landscapes.Nat Commun. 2024 Mar 18;15(1):2370. doi: 10.1038/s41467-024-45909-5. Nat Commun. 2024. PMID: 38499542 Free PMC article.
-
Characterisation of APOBEC3B-Mediated RNA editing in breast cancer cells reveals regulatory roles of NEAT1 and MALAT1 lncRNAs.Oncogene. 2024 Nov;43(46):3366-3377. doi: 10.1038/s41388-024-03171-5. Epub 2024 Sep 25. Oncogene. 2024. PMID: 39322638 Free PMC article.
-
Precise detection of differential RNA editing sites across varied biological conditions using the CADRES pipeline.Sci Rep. 2025 Jun 4;15(1):19683. doi: 10.1038/s41598-025-04957-7. Sci Rep. 2025. PMID: 40467918 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
