

Identification of a Novel Acid Sphingomyelinase Activity Associated with Recombinant Human Acid Ceramidase
- PMID: 38002305
- PMCID: PMC10669851
- DOI: 10.3390/biom13111623
Identification of a Novel Acid Sphingomyelinase Activity Associated with Recombinant Human Acid Ceramidase
Abstract
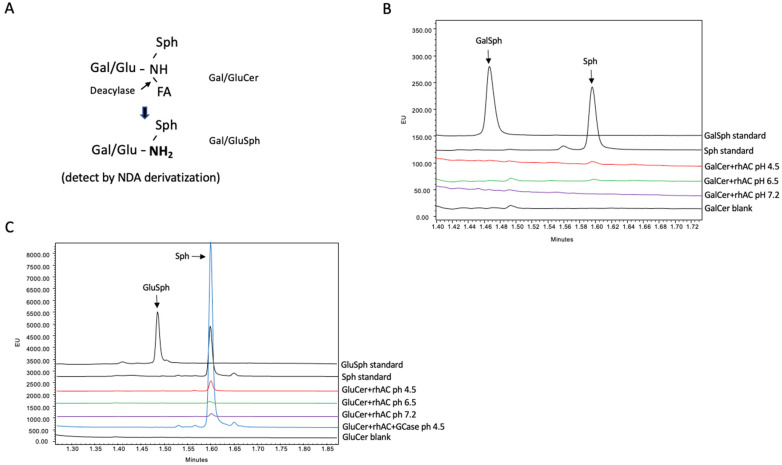
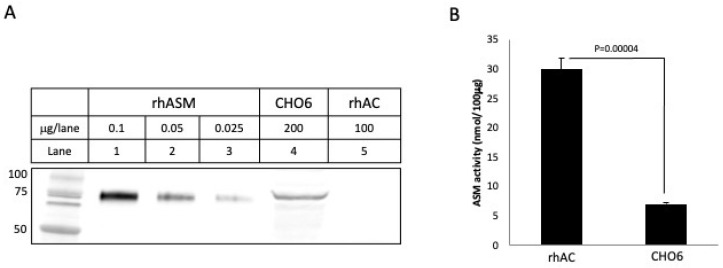
Acid ceramidase (AC) is a lysosomal enzyme required to hydrolyze ceramide to sphingosine by the removal of the fatty acid moiety. An inherited deficiency in this activity results in two disorders, Farber Lipogranulomatosis and spinal muscular atrophy with myoclonic epilepsy, leading to the accumulation of ceramides and other sphingolipids in various cells and tissues. In addition to ceramide hydrolysis, several other activities have been attributed to AC, including a reverse reaction that synthesizes ceramide from free fatty acids and sphingosine, and a deacylase activity that removes fatty acids from complex lipids such as sphingomyelin and glycosphingolipids. A close association of AC with another important enzyme of sphingolipid metabolism, acid sphingomyelinase (ASM), has also been observed. Herein, we used a highly purified recombinant human AC (rhAC) and novel UPLC-based assay methods to investigate the recently described deacylase activity of rhAC against three sphingolipid substrates, sphingomyelin, galactosyl- and glucosylceramide. No deacylase activities were detected using this method, although we did unexpectedly identify a significant ASM activity using natural (C-18) and artificial (Bodipy-C12) sphingomyelin substrates as well as the ASM-specific fluorogenic substrate, hexadecanoylamino-4-methylumbelliferyl phosphorylcholine (HMU-PC). We showed that this ASM activity was not due to contaminating, hamster-derived ASM in the rhAC preparation, and that the treatment of ASM-knockout mice with rhAC significantly reduced sphingomyelin storage in the liver. However, unlike the treatment with rhASM, this did not lead to elevated ceramide or sphingosine levels.
Keywords: Farber disease; ceramidase; deacylase; sphingomyelinase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Deregulation of sphingolipid metabolism in Alzheimer's disease.Neurobiol Aging. 2010 Mar;31(3):398-408. doi: 10.1016/j.neurobiolaging.2008.05.010. Epub 2008 Jun 10. Neurobiol Aging. 2010. PMID: 18547682 Free PMC article.
-
Acid Sphingomyelinase Deficiency Ameliorates Farber Disease.Int J Mol Sci. 2019 Dec 11;20(24):6253. doi: 10.3390/ijms20246253. Int J Mol Sci. 2019. PMID: 31835809 Free PMC article.
-
The reverse activity of human acid ceramidase.J Biol Chem. 2003 Aug 8;278(32):29948-53. doi: 10.1074/jbc.M303310200. Epub 2003 May 22. J Biol Chem. 2003. PMID: 12764132
-
The molecular medicine of acid ceramidase.Biol Chem. 2015 Jun;396(6-7):759-65. doi: 10.1515/hsz-2014-0290. Biol Chem. 2015. PMID: 25938220 Review.
-
Sphingolipids in psychiatric disorders and pain syndromes.Handb Exp Pharmacol. 2013;(216):431-56. doi: 10.1007/978-3-7091-1511-4_22. Handb Exp Pharmacol. 2013. PMID: 23563670 Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
