

Dithiothreitol Affects the Fertilization Response in Immature and Maturing Starfish Oocytes
- PMID: 38002342
- PMCID: PMC10669828
- DOI: 10.3390/biom13111659
Dithiothreitol Affects the Fertilization Response in Immature and Maturing Starfish Oocytes
Abstract
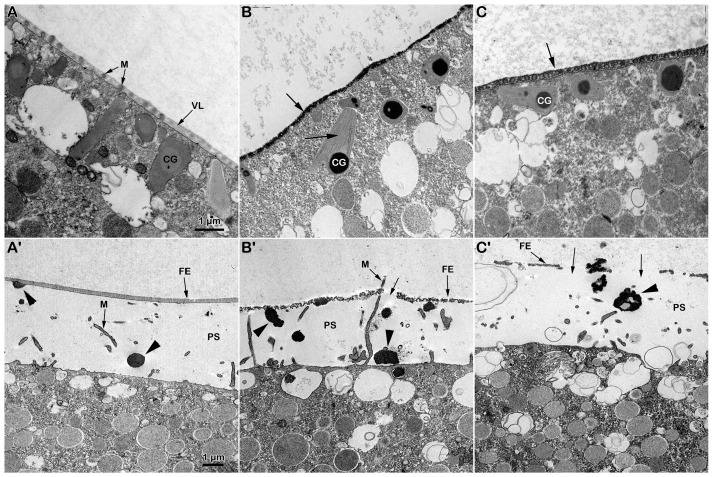
Immature starfish oocytes isolated from the ovary are susceptible to polyspermy due to the structural organization of the vitelline layer covering the oocyte plasma membrane, as well as the distribution and biochemical properties of the actin cytoskeleton of the oocyte cortex. After the resumption of the meiotic cycle of the oocyte triggered by the hormone 1-methyladenine, the maturing oocyte reaches fertilizable conditions to be stimulated by only one sperm with a normal Ca2+ response and cortical reaction. This cytoplasmic ripening of the oocyte, resulting in normal fertilization and development, is due to the remodeling of the cortical actin cytoskeleton and germinal vesicle breakdown (GVBD). Since disulfide-reducing agents such as dithiothreitol (DTT) are known to induce the maturation and GVBD of oocytes in many species of starfish, we analyzed the pattern of the fertilization response displayed by Astropecten aranciacus oocytes pre-exposed to DTT with or without 1-MA stimulation. Short treatment of A. aranciacus immature oocytes with DTT reduced the rate of polyspermic fertilization and altered the sperm-induced Ca2+ response by changing the morphology of microvilli, cortical granules, and biochemical properties of the cortical F-actin. At variance with 1-MA, the DTT treatment of immature starfish oocytes for 70 min did not induce GVBD. On the other hand, the DTT treatment caused an alteration in microvilli morphology and a drastic depolymerization of the cortical F-actin, which impaired the sperm-induced Ca2+ response at fertilization and the subsequent embryonic development.
Keywords: DTT; actin; calcium; disulfide-reducing agents; fertilization; oocyte maturation; polyspermy; starfish.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







Similar articles
-
The Effect of Acidic and Alkaline Seawater on the F-Actin-Dependent Ca2+ Signals Following Insemination of Immature Starfish Oocytes and Mature Eggs.Cells. 2023 Feb 25;12(5):740. doi: 10.3390/cells12050740. Cells. 2023. PMID: 36899875 Free PMC article.
-
Altered actin cytoskeleton in ageing eggs of starfish affects fertilization process.Exp Cell Res. 2019 Aug 15;381(2):179-190. doi: 10.1016/j.yexcr.2019.05.007. Epub 2019 May 10. Exp Cell Res. 2019. PMID: 31082375
-
Cortical changes in starfish (Asterina pectinifera) oocytes during 1-methyladenine-induced maturation and fertilisation/activation.Zygote. 1995 Aug;3(3):225-39. doi: 10.1017/s0967199400002628. Zygote. 1995. PMID: 8903792
-
Maturation and fertilization of echinoderm eggs: Role of actin cytoskeleton dynamics.Biochem Biophys Res Commun. 2018 Nov 25;506(2):361-371. doi: 10.1016/j.bbrc.2018.09.084. Epub 2018 Oct 6. Biochem Biophys Res Commun. 2018. PMID: 30297103 Review.
-
Cellular and molecular aspects of oocyte maturation and fertilization: a perspective from the actin cytoskeleton.Zoological Lett. 2020 Apr 15;6:5. doi: 10.1186/s40851-020-00157-5. eCollection 2020. Zoological Lett. 2020. PMID: 32313685 Free PMC article. Review.
Cited by
-
The Effect of Synthetic Polyamine BPA-C8 on the Fertilization Process of Intact and Denuded Sea Urchin Eggs.Cells. 2024 Sep 2;13(17):1477. doi: 10.3390/cells13171477. Cells. 2024. PMID: 39273047 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
