

Essential Role of Granulosa Cell Glucose and Lipid Metabolism on Oocytes and the Potential Metabolic Imbalance in Polycystic Ovary Syndrome
- PMID: 38003436
- PMCID: PMC10671516
- DOI: 10.3390/ijms242216247
Essential Role of Granulosa Cell Glucose and Lipid Metabolism on Oocytes and the Potential Metabolic Imbalance in Polycystic Ovary Syndrome
Abstract
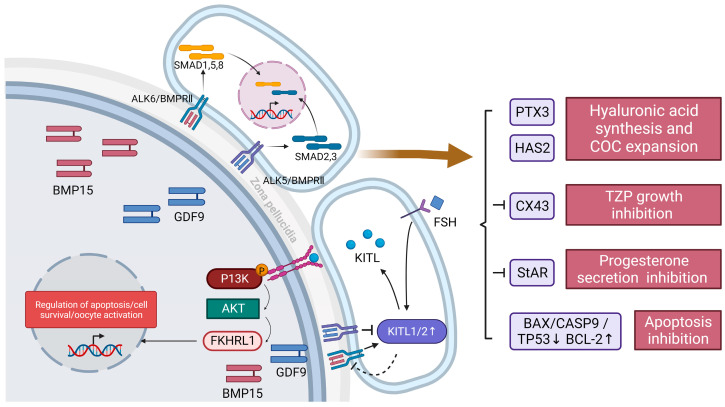
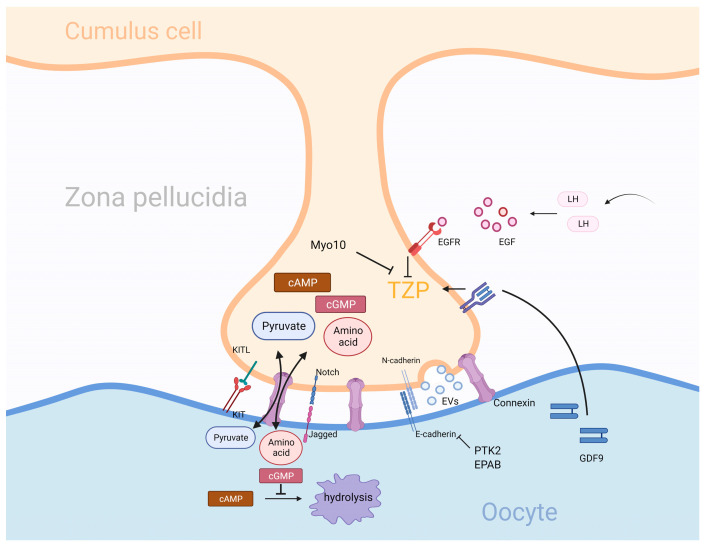
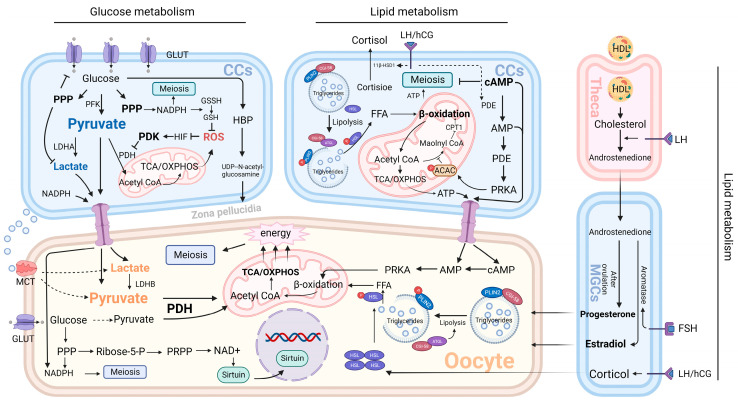
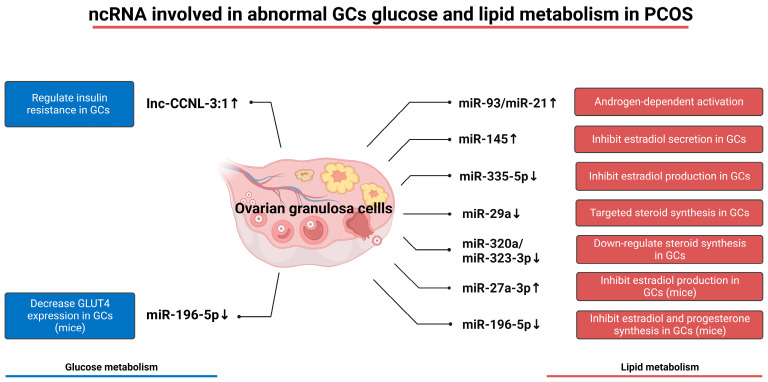
Granulosa cells are crucial for the establishment and maintenance of bidirectional communication among oocytes. Various intercellular material exchange modes, including paracrine and gap junction, are used between them to achieve the efficient delivery of granulosa cell structural components, energy substrates, and signaling molecules to oocytes. Glucose metabolism and lipid metabolism are two basic energy metabolism pathways in granulosa cells; these are involved in the normal development of oocytes. Pyruvate, produced by granulosa cell glycolysis, is an important energy substrate for oocyte development. Granulosa cells regulate changes in intrafollicular hormone levels through the processing of steroid hormones to control the development process of oocytes. This article reviews the material exchange between oocytes and granulosa cells and expounds the significance of granulosa cells in the development of oocytes through both glucose metabolism and lipid metabolism. In addition, we discuss the effects of glucose and lipid metabolism on oocytes under pathological conditions and explore its relationship to polycystic ovary syndrome (PCOS). A series of changes were found in the endogenous molecules and ncRNAs that are related to glucose and lipid metabolism in granulosa cells under PCOS conditions. These findings provide a new therapeutic target for patients with PCOS; additionally, there is potential for improving the fertility of patients with PCOS and the clinical outcomes of assisted reproduction.
Keywords: PCOS; glucose metabolism; granulosa cells; lipid metabolism; oocytes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Mitochondrial and glucose metabolic dysfunctions in granulosa cells induce impaired oocytes of polycystic ovary syndrome through Sirtuin 3.Free Radic Biol Med. 2022 Jul;187:1-16. doi: 10.1016/j.freeradbiomed.2022.05.010. Epub 2022 May 17. Free Radic Biol Med. 2022. PMID: 35594990
-
The effect of metformin treatment in vivo on acute and long-term energy metabolism and progesterone production in vitro by granulosa cells from women with polycystic ovary syndrome.Hum Reprod. 2014 Oct 10;29(10):2302-16. doi: 10.1093/humrep/deu187. Epub 2014 Aug 19. Hum Reprod. 2014. PMID: 25139174 Free PMC article.
-
Different expression and localization of aquaporin 7 and aquaporin 9 in granulosa cells, oocytes, and embryos of patients with polycystic ovary syndrome and the negatively correlated relationship with insulin regulation.Fertil Steril. 2021 Feb;115(2):463-473. doi: 10.1016/j.fertnstert.2020.08.015. Fertil Steril. 2021. PMID: 33579525 Clinical Trial.
-
The Roles of Autophagy in the Genesis and Development of Polycystic Ovary Syndrome.Reprod Sci. 2023 Oct;30(10):2920-2931. doi: 10.1007/s43032-023-01255-3. Epub 2023 May 19. Reprod Sci. 2023. PMID: 37204635 Review.
-
Oxidative stress and energy metabolism abnormalities in polycystic ovary syndrome: from mechanisms to therapeutic strategies.Reprod Biol Endocrinol. 2024 Dec 26;22(1):159. doi: 10.1186/s12958-024-01337-0. Reprod Biol Endocrinol. 2024. PMID: 39722030 Free PMC article. Review.
Cited by
-
Application of amphiregulin in IVM culture of immature human oocytes and pre-insemination culture for COCs in IVF cycles.Front Endocrinol (Lausanne). 2024 Jun 18;15:1428147. doi: 10.3389/fendo.2024.1428147. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 38957445 Free PMC article.
-
The role of mitochondrial dysfunction in ovarian granulosa cells in polycystic ovary syndrome.Endocr Connect. 2025 Jul 12;14(7):e250186. doi: 10.1530/EC-25-0186. Print 2025 Jul 1. Endocr Connect. 2025. PMID: 40586348 Free PMC article. Review.
-
miR-17-5p-Mediated RNA Activation Upregulates KPNA2 Expression and Inhibits High-Glucose-Induced Apoptosis of Sheep Granulosa Cells.Int J Mol Sci. 2025 Jan 23;26(3):943. doi: 10.3390/ijms26030943. Int J Mol Sci. 2025. PMID: 39940713 Free PMC article.
-
Identification and Validation of Genes Involved in Energy Metabolism in Polycystic Ovary Syndrome.Iran J Biotechnol. 2025 Apr 1;23(2):e4102. doi: 10.30498/ijb.2025.514574.4102. eCollection 2025 Apr. Iran J Biotechnol. 2025. PMID: 40860051 Free PMC article.
-
Follicular fluid-derived extracellular vesicles miR-34a-5p regulates granulosa cell glycolysis in polycystic ovary syndrome by targeting LDHA.J Ovarian Res. 2024 Nov 13;17(1):223. doi: 10.1186/s13048-024-01542-w. J Ovarian Res. 2024. PMID: 39538292 Free PMC article.
References
-
- Da Silva-Buttkus P., Jayasooriya G.S., Mora J.M., Mobberley M., Ryder T.A., Baithun M., Stark J., Franks S., Hardy K. Effect of cell shape and packing density on granulosa cell proliferation and formation of multiple layers during early follicle development in the ovary. J. Cell Sci. 2008;121:3890–3900. doi: 10.1242/jcs.036400. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
