

Characterization of VvmiR166s-Target Modules and Their Interaction Pathways in Modulation of Gibberellic-Acid-Induced Grape Seedless Berries
- PMID: 38003470
- PMCID: PMC10670991
- DOI: 10.3390/ijms242216279
Characterization of VvmiR166s-Target Modules and Their Interaction Pathways in Modulation of Gibberellic-Acid-Induced Grape Seedless Berries
Abstract
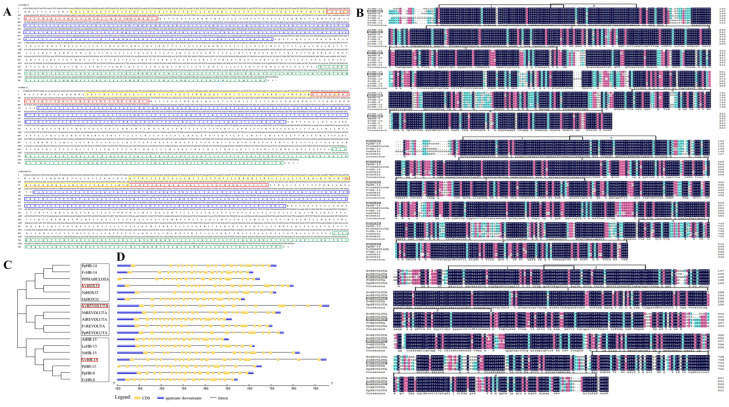
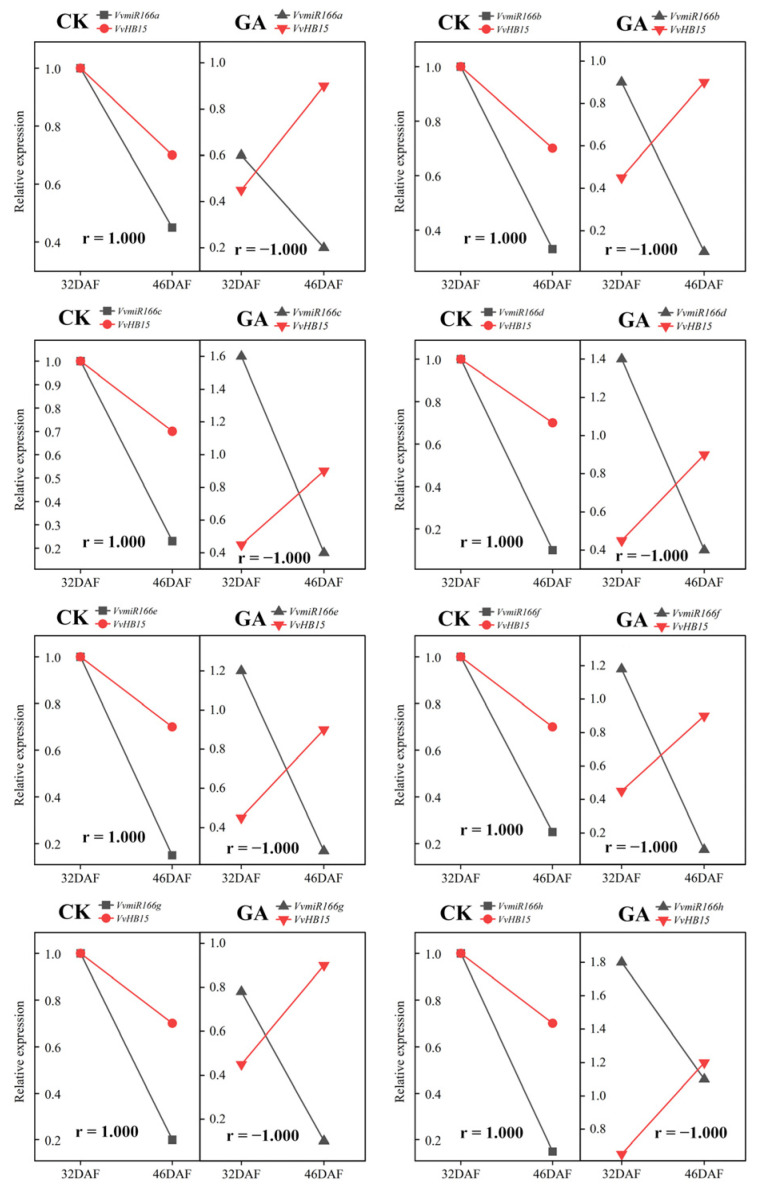
Exogenous GA is widely used to efficiently induce grape seedless berry development for significantly improving berry quality. Recently, we found that VvmiR166s are important regulators of response to GA in grapes, but its roles in GA-induced seedless grape berry development remain elusive. Here, the precise sequences of VvmiR166s and their targets VvREV, VvHB15 and VvHOX32 were determined in grape cv. 'Rosario Bianco', and the cleavage interactions of VvmiR166s-VvHB15/VvHOX32/VvREV modules and the variations in their cleavage roles were confirmed in grape berries. Exogenous GA treatment significantly induced a change in their expression correlations from positive to negative between VvmiR166s and their target genes at the seeds during the stone-hardening stages (32 DAF-46 DAF) in grape berries, indicating exogenous GA change action modes of VvmiR166s on their targets in this process, in which exogenous GA mainly enhanced the negative regulatory roles of VvmiR166s on VvHB15 among all three VvmiR166s-target pairs. The transient OE-VvmiR166a-h/OE-VvHB15 in tobacco confirmed that out of the VvmiR166 family, VvmiR166h/a/b might be the main factors in modulating lignin synthesis through inhibiting VvHB15, of which VvmiR166h-VvHB15-NtPAL4/NtCCR1/NtCCR2/NtCCoAMT5/NtCOMT1 and VvmiR166a/b-VvHB15-NtCAD1 are the potential key regulatory modules in lignin synthesis. Together with the GA-induced expression modes of VvmiR166s-VvHB15 and genes related to lignin synthesis in grape berries, we revealed that GA might repress lignin synthesis mainly by repressing VvCAD1/VvCCR2/VvPAL2/VvPAL3/Vv4CL/VvLac7 levels via mediating VvmiR166s-VvHB15 modules in GA-induced grape seedless berries. Our findings present a novel insight into the roles of VvmiR66s that are responsive to GA in repressing the lignin synthesis of grape seedless berries, with different lignin-synthesis-enzyme-dependent action pathways in diverse plants, which have important implications for the molecular breeding of high-quality seedless grape berries.
Keywords: HD-Zip III; VvmiR166s; gibberellin; lignin; seedless grape.
Conflict of interest statement
The authors declare no conflict of interest.
Figures











Similar articles
-
Gibberellin Mediates VvmiR397a-VvLAC4 via VvSLR1-VvWRKY26 Cascade Signal to Repress the Seed-Stone Development During GA-Induced Grape Parthenocarpy.Plant Cell Environ. 2025 Aug;48(8):5930-5949. doi: 10.1111/pce.15570. Epub 2025 Apr 23. Plant Cell Environ. 2025. PMID: 40269609
-
Identification of miRNAs-mediated seed and stone-hardening regulatory networks and their signal pathway of GA-induced seedless berries in grapevine (V. vinifera L.).BMC Plant Biol. 2021 Sep 29;21(1):442. doi: 10.1186/s12870-021-03188-y. BMC Plant Biol. 2021. PMID: 34587914 Free PMC article.
-
VvmiR160s/VvARFs interaction and their spatio-temporal expression/cleavage products during GA-induced grape parthenocarpy.BMC Plant Biol. 2019 Mar 21;19(1):111. doi: 10.1186/s12870-019-1719-9. BMC Plant Biol. 2019. PMID: 30898085 Free PMC article.
-
miR3633a-GA3ox2 Module Conducts Grape Seed-Embryo Abortion in Response to Gibberellin.Int J Mol Sci. 2022 Aug 7;23(15):8767. doi: 10.3390/ijms23158767. Int J Mol Sci. 2022. PMID: 35955901 Free PMC article.
-
Identification and Characterization of AUXIN Response Factor Gene Family Reveals Their Regulatory Network to Respond the Multi-Hormones Crosstalk during GA-Induced Grape Parthenocarpic Berry.Int J Mol Sci. 2022 Sep 21;23(19):11108. doi: 10.3390/ijms231911108. Int J Mol Sci. 2022. PMID: 36232409 Free PMC article.
References
-
- Acheampong A.K., Hu J., Rotman A., Zheng C., Halaly T., Takebayashi Y., Jikumaru Y., Kamiya Y., Lichter A., Sun T.P., et al. Functional characterization and developmental expression profiling of gibberellin signalling components in Vitis vinifera. J. Exp. Bot. 2015;66:1463–1476. doi: 10.1093/jxb/eru504. - DOI - PMC - PubMed
-
- Ikeda F., Ishikawa K., Yazawa S., Baba T. Induction of compact clusters with large seedless berries in the grape cultivar ’Fujiminori’ by the use of streptomycin, gibberellins, and CPPU. Vitic.-Living Limit. 2004;640:361–368. doi: 10.17660/ActaHortic.2004.640.43. - DOI
-
- Fujishima H., Matsuda K., Ushijima K., Yahata D., Shiraishi M., Chijiwa H. Differences in Fruit Quality between Gibberellin-Treated and Untreated Berries of ‘Kyoho’ Grapes. Hortic. Res. 2012;11:405–410. doi: 10.2503/hrj.11.405. - DOI
MeSH terms
Substances
Grants and funding
- 31972373/National Natural Science Funds
- JBGS [2021]086/Jiangsu province seed industry revitalization of the leading project
- BK20181318, BK20160587/Jiangsu Province Natural Science Funds
- 2018YFD1000106/the National Key Research and Development Program of China
- KYTZ201602/Central University basic scientific research business fee independent innovation major project (NATURAL SCIENCE)
LinkOut - more resources
Full Text Sources
Miscellaneous
