

Transcriptome Profiling Provides Insights into the Early Development of Tiller Buds in High- and Low-Tillering Orchardgrass Genotypes
- PMID: 38003564
- PMCID: PMC10671593
- DOI: 10.3390/ijms242216370
Transcriptome Profiling Provides Insights into the Early Development of Tiller Buds in High- and Low-Tillering Orchardgrass Genotypes
Abstract
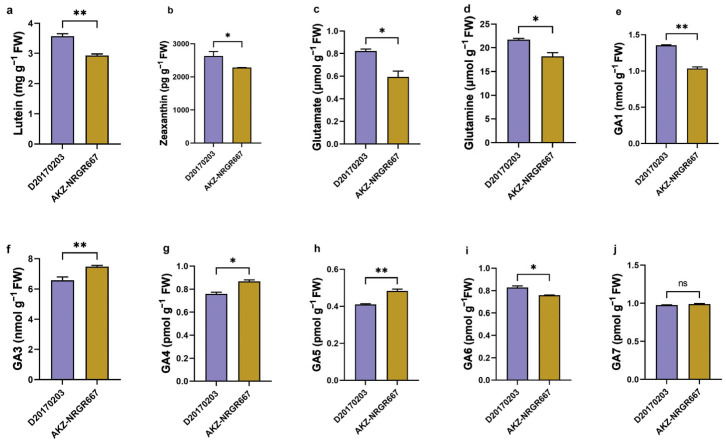
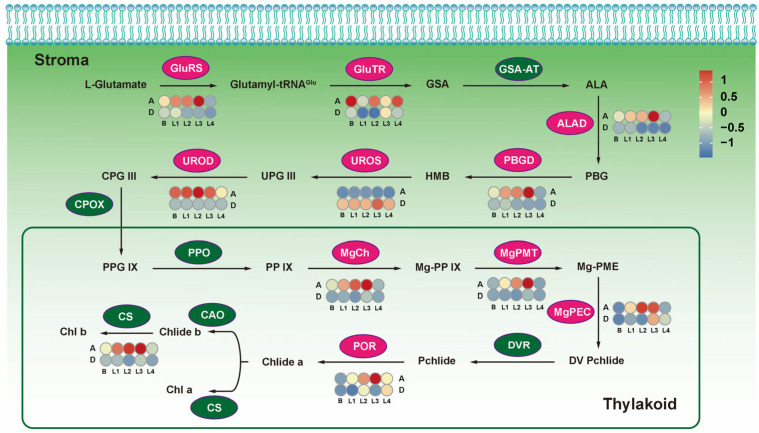
Orchardgrass (Dactylis glomerata L.) is among the most economically important perennial cool-season grasses, and is considered an excellent hay, pasture, and silage crop in temperate regions worldwide. Tillering is a vital feature that dominates orchardgrass regeneration and biomass yield. However, transcriptional dynamics underlying early-stage bud development in high- and low-tillering orchardgrass genotypes are unclear. Thus, this study assessed the photosynthetic parameters, the partially essential intermediate biomolecular substances, and the transcriptome to elaborate the early-stage profiles of tiller development. Photosynthetic efficiency and morphological development significantly differed between high- (AKZ-NRGR667) and low-tillering genotypes (D20170203) at the early stage after tiller formation. The 206.41 Gb of high-quality reads revealed stage-specific differentially expressed genes (DEGs), demonstrating that signal transduction and energy-related metabolism pathways, especially photosynthetic-related processes, influence tiller induction and development. Moreover, weighted correlation network analysis (WGCNA) and functional enrichment identified distinctively co-expressed gene clusters and four main regulatory pathways, including chlorophyll, lutein, nitrogen, and gibberellic acid (GA) metabolism pathways. Therefore, photosynthesis, carbohydrate synthesis, nitrogen efficient utilization, and phytohormone signaling pathways are closely and intrinsically linked at the transcriptional level. These findings enhance our understanding of tillering in orchardgrass and perennial grasses, providing a new breeding strategy for improving forage biomass yield.
Keywords: orchardgrass; perennial forage; photosynthesis; tillering regulation; transcriptome profiling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







Similar articles
-
Comparative transcriptome analyses reveal different mechanism of high- and low-tillering genotypes controlling tiller growth in orchardgrass (Dactylis glomerata L.).BMC Plant Biol. 2020 Aug 5;20(1):369. doi: 10.1186/s12870-020-02582-2. BMC Plant Biol. 2020. PMID: 32758131 Free PMC article.
-
Transcriptome Analysis Reveals the Potential Molecular Mechanisms of Tiller Bud Development in Orchardgrass.Int J Mol Sci. 2023 Oct 30;24(21):15762. doi: 10.3390/ijms242115762. Int J Mol Sci. 2023. PMID: 37958746 Free PMC article.
-
Analysis of controlling genes for tiller growth of Psathyrostachys juncea based on transcriptome sequencing technology.BMC Plant Biol. 2022 Sep 23;22(1):456. doi: 10.1186/s12870-022-03837-w. BMC Plant Biol. 2022. PMID: 36151542 Free PMC article.
-
Genetic and molecular insights into tiller development and approaches for crop yield improvement.Front Plant Sci. 2025 Mar 27;16:1532180. doi: 10.3389/fpls.2025.1532180. eCollection 2025. Front Plant Sci. 2025. PMID: 40212868 Free PMC article. Review.
-
Genetic Basis of Tillering Angle from Other Plants to Wheat: Current Progress and Future Perspectives.Plants (Basel). 2024 Nov 18;13(22):3237. doi: 10.3390/plants13223237. Plants (Basel). 2024. PMID: 39599446 Free PMC article. Review.
Cited by
-
Genetic Mapping by 55K Single-Nucleotide Polymorphism Array Reveals Candidate Genes for Tillering Trait in Wheat Mutant dmc.Genes (Basel). 2024 Dec 22;15(12):1652. doi: 10.3390/genes15121652. Genes (Basel). 2024. PMID: 39766919 Free PMC article.
References
-
- Feng G., Xu L., Wang J., Nie G., Bushman B.S., Xie W., Yan H., Yang Z., Guan H., Huang L., et al. Integration of small RNAs and transcriptome sequencing uncovers a complex regulatory network during vernalization and heading stages of orchardgrass (Dactylis glomerata L.) BMC Genom. 2018;19:727. doi: 10.1186/s12864-018-5104-0. - DOI - PMC - PubMed
-
- Xie K., Cakmak I., Wang S., Zhang F., Guo S. Synergistic and antagonistic interactions between potassium and magnesium in higher plants. Crop J. 2021;9:249–256. doi: 10.1016/j.cj.2020.10.005. - DOI
-
- Rotili D.H., Sadras V.O., Abeledo L.G., Ferreyra J.M., Micheloud J.R., Duarte G., Girón P., Ermácora M., Maddonni G.A. Impacts of vegetative and reproductive plasticity associated with tillering in maize crops in low-yielding environments: A physiological framework. Field Crops Res. 2021;265:108107. doi: 10.1016/j.fcr.2021.108107. - DOI
MeSH terms
Substances
Grants and funding
- 2021YFYZ0013/the Forage Breeding Project of Sichuan Province
- No. CARS-34/the Modern Agro-industry Technology Research System
- NSFC 32101422/the National Natural Science Foundation of China
- cstc2021jcyj-msxmX0865/the Natural Science Foundation of Chongqing
- 2022YFH0058/the Sichuan Province's Science Fund for International Cooperation
LinkOut - more resources
Full Text Sources
