

ZBP1 Drives IAV-Induced NLRP3 Inflammasome Activation and Lytic Cell Death, PANoptosis, Independent of the Necroptosis Executioner MLKL
- PMID: 38005819
- PMCID: PMC10674287
- DOI: 10.3390/v15112141
ZBP1 Drives IAV-Induced NLRP3 Inflammasome Activation and Lytic Cell Death, PANoptosis, Independent of the Necroptosis Executioner MLKL
Abstract
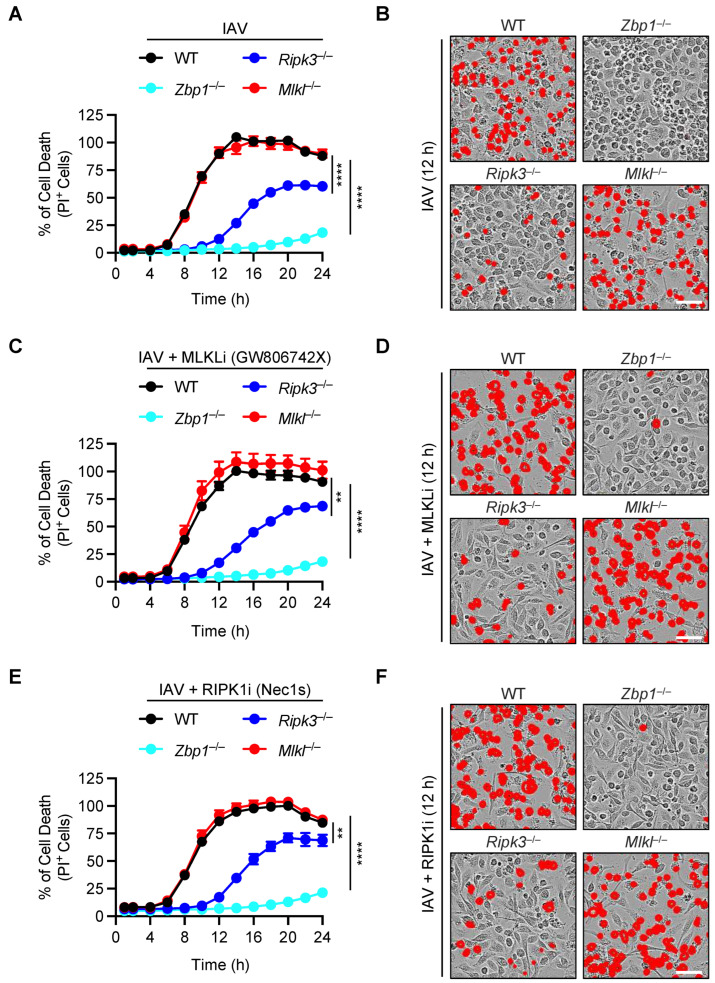
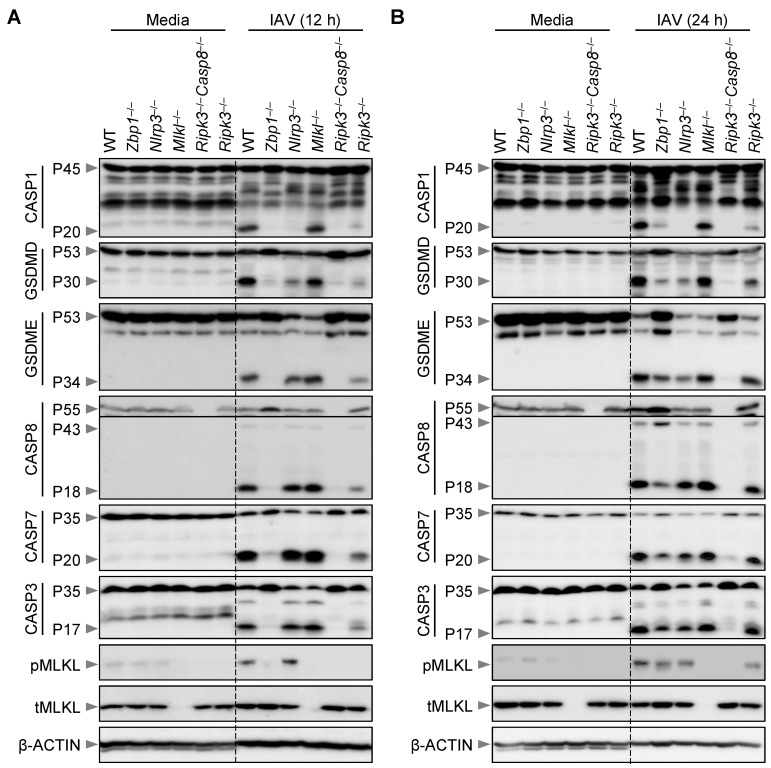
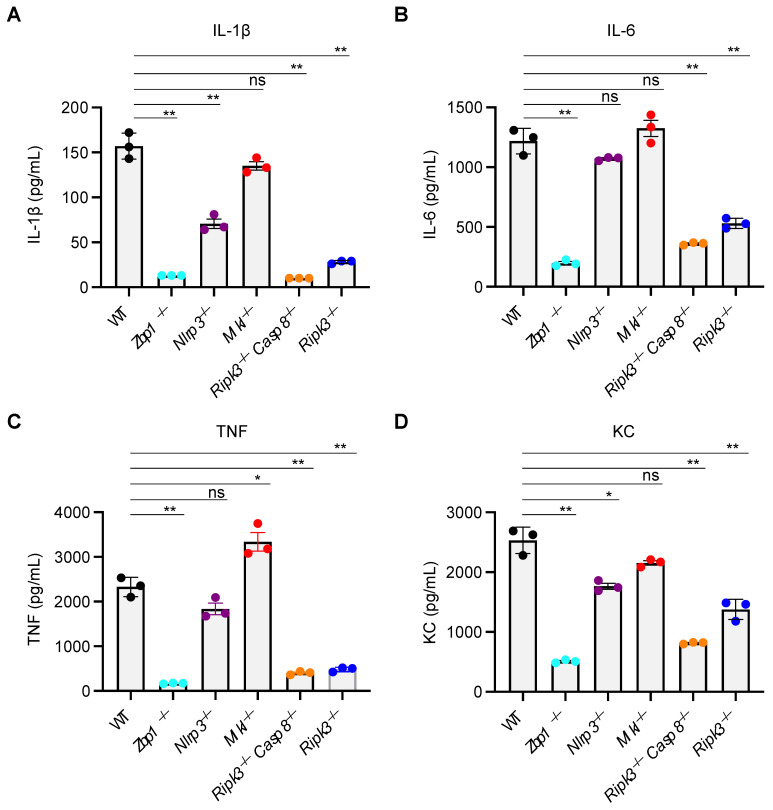
Influenza A virus (IAV) continues to pose a significant global health threat, causing severe respiratory infections that result in substantial annual morbidity and mortality. Recent research highlights the pivotal role of innate immunity, cell death, and inflammation in exacerbating the severity of respiratory viral diseases. One key molecule in this process is ZBP1, a well-recognized innate immune sensor for IAV infection. Upon activation, ZBP1 triggers the formation of a PANoptosome complex containing ASC, caspase-8, and RIPK3, among other molecules, leading to inflammatory cell death, PANoptosis, and NLRP3 inflammasome activation for the maturation of IL-1β and IL-18. However, the role for other molecules in this process requires further evaluation. In this study, we investigated the role of MLKL in regulating IAV-induced cell death and NLRP3 inflammasome activation. Our data indicate IAV induced inflammatory cell death through the ZBP1-PANoptosome, where caspases and RIPKs serve as core components. However, IAV-induced lytic cell death was only partially dependent on RIPK3 at later timepoints and was fully independent of MLKL throughout all timepoints tested. Additionally, NLRP3 inflammasome activation was unaffected in MLKL-deficient cells, establishing that MLKL and MLKL-dependent necroptosis do not act upstream of NLRP3 inflammasome activation, IL-1β maturation, and lytic cell death during IAV infection.
Keywords: IAV; MLKL; NLRP3; PANoptosis; RIPK3; apoptosis; caspase; cell death; executioner; gasdermin; inflammasome; necroptosis; pyroptosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
MLKL-Driven Inflammasome Activation and Caspase-8 Mediate Inflammatory Cell Death in Influenza A Virus Infection.mBio. 2023 Apr 25;14(2):e0011023. doi: 10.1128/mbio.00110-23. Epub 2023 Feb 28. mBio. 2023. PMID: 36852999 Free PMC article.
-
The regulation of the ZBP1-NLRP3 inflammasome and its implications in pyroptosis, apoptosis, and necroptosis (PANoptosis).Immunol Rev. 2020 Sep;297(1):26-38. doi: 10.1111/imr.12909. Epub 2020 Jul 29. Immunol Rev. 2020. PMID: 32729116 Free PMC article. Review.
-
Innate immune sensor NLRP3 drives PANoptosome formation and PANoptosis.J Immunol. 2025 Jun 1;214(6):1236-1246. doi: 10.1093/jimmun/vkaf042. J Immunol. 2025. PMID: 40249072 Free PMC article.
-
Immune regulator IRF1 contributes to ZBP1-, AIM2-, RIPK1-, and NLRP12-PANoptosome activation and inflammatory cell death (PANoptosis).J Biol Chem. 2023 Sep;299(9):105141. doi: 10.1016/j.jbc.2023.105141. Epub 2023 Aug 7. J Biol Chem. 2023. PMID: 37557956 Free PMC article.
-
It's All in the PAN: Crosstalk, Plasticity, Redundancies, Switches, and Interconnectedness Encompassed by PANoptosis Underlying the Totality of Cell Death-Associated Biological Effects.Cells. 2022 Apr 29;11(9):1495. doi: 10.3390/cells11091495. Cells. 2022. PMID: 35563804 Free PMC article. Review.
Cited by
-
PANoptosis in autoimmune diseases interplay between apoptosis, necrosis, and pyroptosis.Front Immunol. 2024 Oct 31;15:1502855. doi: 10.3389/fimmu.2024.1502855. eCollection 2024. Front Immunol. 2024. PMID: 39544942 Free PMC article. Review.
-
Trimethylamine Induced Chronic Kidney Injury by Activating the ZBP1-NLRP3 Inflammasome Pathway.Physiol Res. 2024 Nov 15;73(5):779-789. doi: 10.33549/physiolres.935378. Physiol Res. 2024. PMID: 39545792 Free PMC article.
-
TRIF-TAK1 signaling suppresses caspase-8/3-mediated GSDMD/E activation and pyroptosis in influenza A virus-infected airway epithelial cells.iScience. 2024 Dec 12;28(1):111581. doi: 10.1016/j.isci.2024.111581. eCollection 2025 Jan 17. iScience. 2024. PMID: 39811662 Free PMC article.
-
Nd:YAG1064nm laser functions against Sporothrix globosa by inducing PANoptosis via the regulation of ZBP1-induced PANoptosome activation.Front Microbiol. 2025 Mar 26;16:1555338. doi: 10.3389/fmicb.2025.1555338. eCollection 2025. Front Microbiol. 2025. PMID: 40207151 Free PMC article.
-
Age-related STING suppression in macrophages contributes to increased viral load during influenza a virus infection.Immun Ageing. 2024 Nov 14;21(1):80. doi: 10.1186/s12979-024-00482-9. Immun Ageing. 2024. PMID: 39543713 Free PMC article.
References
-
- Iuliano A.D., Roguski K.M., Chang H.H., Muscatello D.J., Palekar R., Tempia S., Cohen C., Gran J.M., Schanzer D., Cowling B.J., et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet. 2018;391:1285–1300. doi: 10.1016/S0140-6736(17)33293-2. - DOI - PMC - PubMed
-
- Kash J.C., Tumpey T.M., Proll S.C., Carter V., Perwitasari O., Thomas M.J., Basler C.F., Palese P., Taubenberger J.K., Garcia-Sastre A., et al. Genomic analysis of increased host immune and cell death responses induced by 1918 influenza virus. Nature. 2006;443:578–581. doi: 10.1038/nature05181. - DOI - PMC - PubMed
-
- Schulert G.S., Zhang M., Fall N., Husami A., Kissell D., Hanosh A., Zhang K., Davis K., Jentzen J.M., Napolitano L., et al. Whole-Exome Sequencing Reveals Mutations in Genes Linked to Hemophagocytic Lymphohistiocytosis and Macrophage Activation Syndrome in Fatal Cases of H1N1 Influenza. J. Infect. Dis. 2016;213:1180–1188. doi: 10.1093/infdis/jiv550. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
