

A Novel Quadruple Gene-Deleted BoHV-1-Vectored RVFV Subunit Vaccine Induces Humoral and Cell-Mediated Immune Response against Rift Valley Fever in Calves
- PMID: 38005861
- PMCID: PMC10674938
- DOI: 10.3390/v15112183
A Novel Quadruple Gene-Deleted BoHV-1-Vectored RVFV Subunit Vaccine Induces Humoral and Cell-Mediated Immune Response against Rift Valley Fever in Calves
Abstract
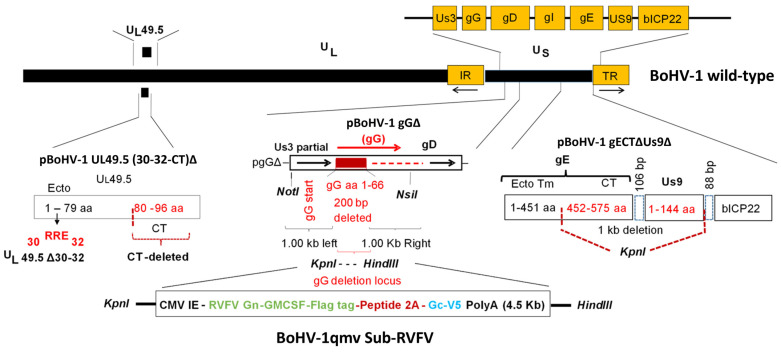
Rift Valley fever virus (RVFV) is considered to be a high biodefense priority based on its threat to livestock and its ability to cause human hemorrhagic fever. RVFV-infected livestock are also a significant risk factor for human infection by direct contact with contaminated blood, tissues, and aborted fetal materials. Therefore, livestock vaccination in the affected regions has the direct dual benefit and one-health approach of protecting the lives of millions of animals and eliminating the risk of severe and sometimes lethal human Rift Valley fever (RVF) disease. Recently, we have developed a bovine herpesvirus type 1 (BoHV-1) quadruple gene mutant virus (BoHV-1qmv) vector that lacks virulence and immunosuppressive properties due to the deletion of envelope proteins UL49.5, glycoprotein G (gG), gE cytoplasmic tail, and US9 coding sequences. In the current study, we engineered the BoHV-1qmv further by incorporating a chimeric gene sequence to express a proteolytically cleavable polyprotein: RVFV envelope proteins Gn ectodomain sequence fused with bovine granulocyte-macrophage colony-stimulating factor (GMCSF) and Gc, resulting in a live BoHV-1qmv-vectored subunit vaccine against RVFV for livestock. In vitro, the resulting recombinant virus, BoHV-1qmv Sub-RVFV, was replicated in cell culture with high titers. The chimeric Gn-GMCSF and Gc proteins expressed by the vaccine virus formed the Gn-Gc complex. In calves, the BoHV-1qmv Sub-RVFV vaccination was safe and induced moderate levels of the RVFV vaccine strain, MP12-specific neutralizing antibody titers. Additionally, the peripheral blood mononuclear cells from the vaccinated calves had six-fold increased levels of interferon-gamma transcription compared with that of the BoHV-1qmv (vector)-vaccinated calves when stimulated with heat-inactivated MP12 antigen in vitro. Based on these findings, we believe that a single dose of BoHV-1qmv Sub-RVFV vaccine generated a protective RVFV-MP12-specific humoral and cellular immune response. Therefore, the BoHV-1qmv sub-RVFV can potentially be a protective subunit vaccine for cattle against RVFV.
Keywords: BoHV-1 mutant; BoHV-1 vector; Gn and Gc; RVFV; cattle; immunogenicity; subunit-vaccine.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
 ) in immunized calves assessed by qPCR and virus isolation. (A) After the inoculation into the KOP-R cells, the virus was isolated from each animal’s nasal swab and titrated in confluent KOP-R cells by plaque assay. Shown are the virus titers in the plaque-forming unit/mL of the nasal swab. (B) DNA was isolated from nasal swabs following inoculation and BoHV-1-qPCR was performed. The mean copy numbers of the BoHV-1 genome are shown in 100 ng of total DNA. PFU/mL; dpv—days post-vaccination.
) in immunized calves assessed by qPCR and virus isolation. (A) After the inoculation into the KOP-R cells, the virus was isolated from each animal’s nasal swab and titrated in confluent KOP-R cells by plaque assay. Shown are the virus titers in the plaque-forming unit/mL of the nasal swab. (B) DNA was isolated from nasal swabs following inoculation and BoHV-1-qPCR was performed. The mean copy numbers of the BoHV-1 genome are shown in 100 ng of total DNA. PFU/mL; dpv—days post-vaccination.
Similar articles
-
A Novel BoHV-1-Vectored Subunit RVFV Vaccine Induces a Robust Humoral and Cell-Mediated Immune Response Against Rift Valley Fever in Sheep.Viruses. 2025 Feb 23;17(3):304. doi: 10.3390/v17030304. Viruses. 2025. PMID: 40143235 Free PMC article.
-
A Quadruple Gene-Deleted Live BoHV-1 Subunit RVFV Vaccine Vector Reactivates from Latency and Replicates in the TG Neurons of Calves but Is Not Transported to and Shed from Nasal Mucosa.Viruses. 2024 Sep 21;16(9):1497. doi: 10.3390/v16091497. Viruses. 2024. PMID: 39339973 Free PMC article.
-
Vaccination with DNA plasmids expressing Gn coupled to C3d or alphavirus replicons expressing gn protects mice against Rift Valley fever virus.PLoS Negl Trop Dis. 2010 Jun 22;4(6):e725. doi: 10.1371/journal.pntd.0000725. PLoS Negl Trop Dis. 2010. PMID: 20582312 Free PMC article.
-
Single-cycle replicable Rift Valley fever virus mutants as safe vaccine candidates.Virus Res. 2016 May 2;216:55-65. doi: 10.1016/j.virusres.2015.05.012. Epub 2015 May 27. Virus Res. 2016. PMID: 26022573 Free PMC article. Review.
-
Novel approaches to develop Rift Valley fever vaccines.Front Cell Infect Microbiol. 2012 Oct 26;2:131. doi: 10.3389/fcimb.2012.00131. eCollection 2012. Front Cell Infect Microbiol. 2012. PMID: 23112960 Free PMC article. Review.
Cited by
-
The Adaptive Immune Response against Bunyavirales.Viruses. 2024 Mar 21;16(3):483. doi: 10.3390/v16030483. Viruses. 2024. PMID: 38543848 Free PMC article. Review.
-
A Novel BoHV-1-Vectored Subunit RVFV Vaccine Induces a Robust Humoral and Cell-Mediated Immune Response Against Rift Valley Fever in Sheep.Viruses. 2025 Feb 23;17(3):304. doi: 10.3390/v17030304. Viruses. 2025. PMID: 40143235 Free PMC article.
-
A Quadruple Gene-Deleted Live BoHV-1 Subunit RVFV Vaccine Vector Reactivates from Latency and Replicates in the TG Neurons of Calves but Is Not Transported to and Shed from Nasal Mucosa.Viruses. 2024 Sep 21;16(9):1497. doi: 10.3390/v16091497. Viruses. 2024. PMID: 39339973 Free PMC article.
References
-
- Van Velden D.J., Meyer J.D., Olivier J., Gear J.H., McIntosh B. Rift Valley fever affecting humans in South Africa: A clinicopathological study. S. Afr. Med. J. 1977;51:867–871. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
