

Identification of Host Factors for Rift Valley Fever Phlebovirus
- PMID: 38005928
- PMCID: PMC10675714
- DOI: 10.3390/v15112251
Identification of Host Factors for Rift Valley Fever Phlebovirus
Abstract
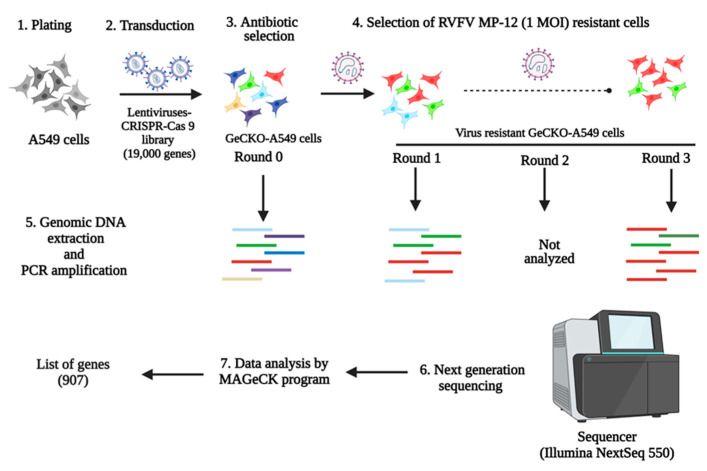
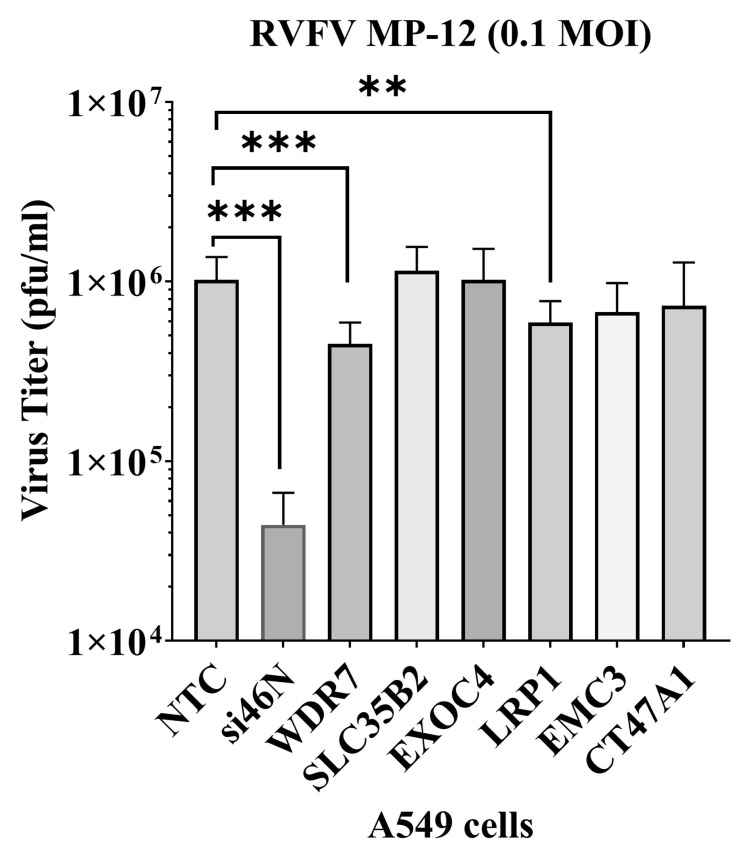
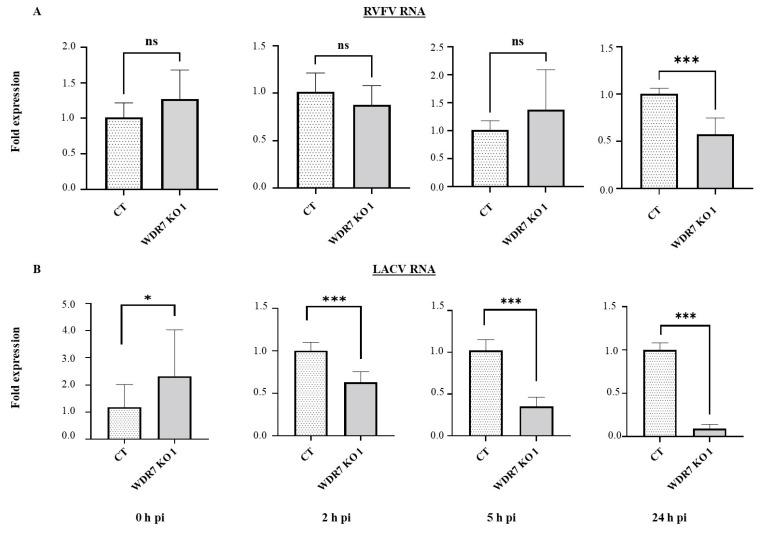
Rift Valley fever phlebovirus (RVFV) is a zoonotic pathogen that causes Rift Valley fever (RVF) in livestock and humans. Currently, there is no licensed human vaccine or antiviral drug to control RVF. Although multiple species of animals and humans are vulnerable to RVFV infection, host factors affecting susceptibility are not well understood. To identify the host factors or genes essential for RVFV replication, we conducted CRISPR-Cas9 knockout screening in human A549 cells. We then validated the putative genes using siRNA-mediated knock-downs and CRISPR-Cas9-mediated knock-out studies. The role of a candidate gene in the virus replication cycle was assessed by measuring intracellular viral RNA accumulation, and the virus titers were analyzed using plaque assay or TCID50 assay. We identified approximately 900 genes with potential involvement in RVFV infection and replication. Further evaluation of the effect of six genes on viral replication using siRNA-mediated knock-downs revealed that silencing two genes (WDR7 and LRP1) significantly impaired RVFV replication. For further analysis, we focused on the WDR7 gene since the role of the LRP1 gene in RVFV replication was previously described in detail. WDR7 knockout A549 cell lines were generated and used to dissect the effect of WRD7 on a bunyavirus, RVFV, and an orthobunyavirus, La Crosse encephalitis virus (LACV). We observed significant effects of WDR7 knockout cells on both intracellular RVFV RNA levels and viral titers. At the intracellular RNA level, WRD7 affected RVFV replication at a later phase of its replication cycle (24 h) when compared with the LACV replication, which was affected in an earlier replication phase (12 h). In summary, we identified WDR7 as an essential host factor for the replication of two different viruses, RVFV and LACV, both of which belong to the Bunyavirales order. Future studies will investigate the mechanistic role through which WDR7 facilitates phlebovirus replication.
Keywords: A549 cells; LACV; MP-12; RVFV; WDR7 gene; bunyavirus; host factor; phlebovirus.
Conflict of interest statement
All authors declare no conflicts of interest. The JAR laboratory received support from Tonix Pharmaceuticals, Xing Technologies, Esperovax, and Zoetis, outside of the reported work. JAR is the inventor of patents and patent applications on the use of antivirals and vaccines for the treatment and prevention of virus infections, owned by Kansas State University, KS.
Figures
Update of
-
Identification of host factors for Rift Valley Fever Phlebovirus.bioRxiv [Preprint]. 2023 Sep 29:2023.09.28.559935. doi: 10.1101/2023.09.28.559935. bioRxiv. 2023. Update in: Viruses. 2023 Nov 13;15(11):2251. doi: 10.3390/v15112251. PMID: 37808812 Free PMC article. Updated. Preprint.
References
-
- Daubney R., Hudson J.R., Garnham P.C. Enzootic hepatitis or rift valley fever. An undescribed virus disease of sheep cattle and man from east africa. J. Pathol. Bacteriol. 1931;34:545–579. doi: 10.1002/path.1700340418. - DOI
-
- Shoemaker T., Boulianne C., Vincent M.J., Pezzanite L., Al-Qahtani M.M., Al-Mazrou Y., Khan A.S., Rollin P.E., Swanepoel R., Ksiazek T.G., et al. Genetic analysis of viruses associated with emergence of Rift Valley fever in Saudi Arabia and Yemen, 2000–2001. Emerg. Infect. Dis. 2002;8:1415–1420. doi: 10.3201/eid0812.020195. - DOI - PMC - PubMed
-
- Coetzer J.A. Natural Cases in New-Born Lambs. Onderstepoort J.Vet.Res. 1977;44:205–211. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
