

Characterization of the Efficacy of a Split Swine Influenza A Virus Nasal Vaccine Formulated with a Nanoparticle/STING Agonist Combination Adjuvant in Conventional Pigs
- PMID: 38006039
- PMCID: PMC10675483
- DOI: 10.3390/vaccines11111707
Characterization of the Efficacy of a Split Swine Influenza A Virus Nasal Vaccine Formulated with a Nanoparticle/STING Agonist Combination Adjuvant in Conventional Pigs
Abstract
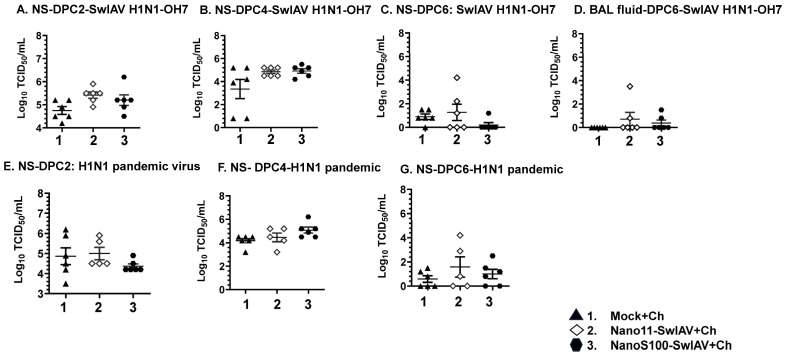
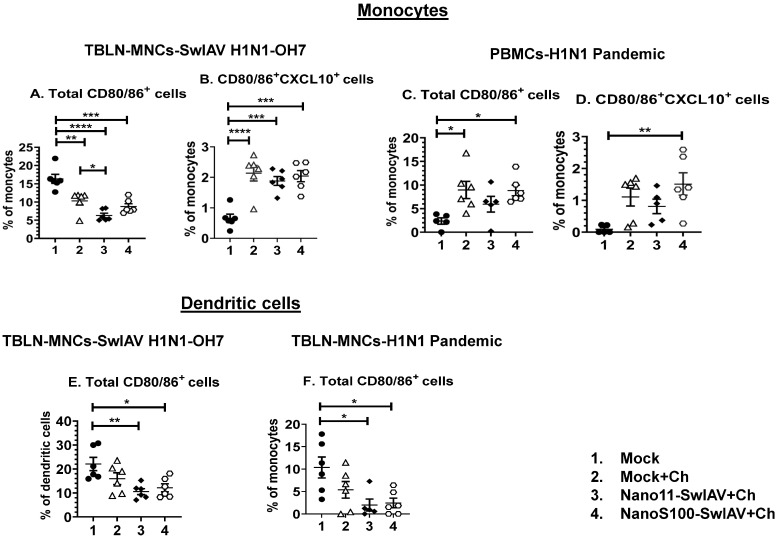
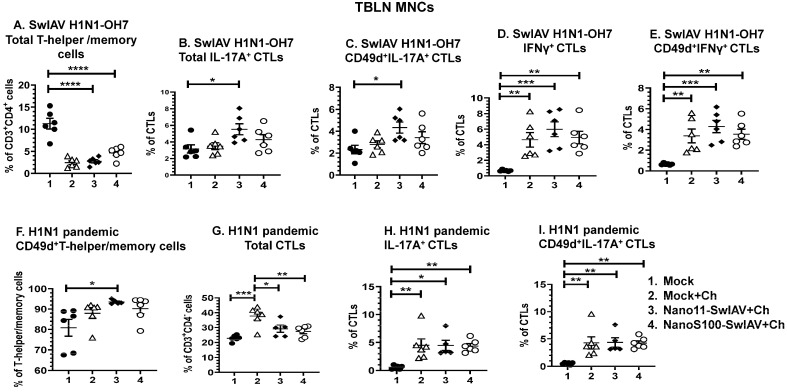
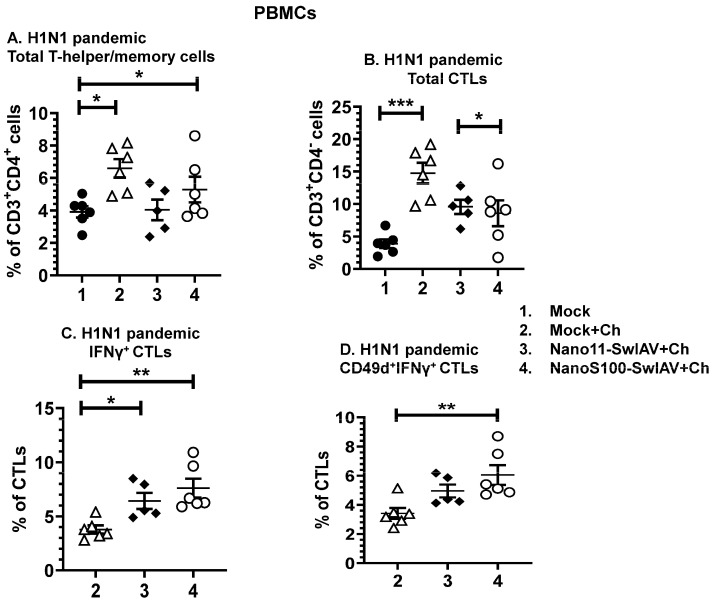
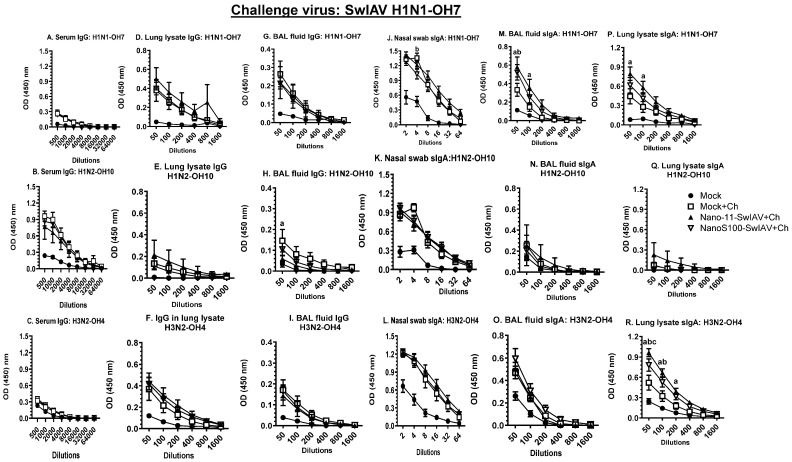
Swine influenza A viruses (SwIAVs) are pathogens of both veterinary and medical significance. Intranasal (IN) vaccination has the potential to reduce flu infection. We investigated the efficacy of split SwIAV H1N2 antigens adsorbed with a plant origin nanoparticle adjuvant [Nano11-SwIAV] or in combination with a STING agonist ADU-S100 [NanoS100-SwIAV]. Conventional pigs were vaccinated via IN and challenged with a heterologous SwIAV H1N1-OH7 or 2009 H1N1 pandemic virus. Immunologically, in NanoS100-SwIAV vaccinates, we observed enhanced frequencies of activated monocytes in the blood of the pandemic virus challenged animals and in tracheobronchial lymph nodes (TBLN) of H1N1-OH7 challenged animals. In both groups of the virus challenged pigs, increased frequencies of IL-17A+ and CD49d+IL-17A+ cytotoxic lymphocytes were observed in Nano11-SwIAV vaccinates in the draining TBLN. Enhanced frequency of CD49d+IFNγ+ CTLs in the TBLN and blood of both the Nano11-based SwIAV vaccinates was observed. Animals vaccinated with both Nano11-based vaccines had upregulated cross-reactive secretory IgA in the lungs and serum IgG against heterologous and heterosubtypic viruses. However, in NanoS100-SwIAV vaccinates, a slight early reduction in the H1N1 pandemic virus and a late reduction in the SwIAV H1N1-OH7 load in the nasal passages were detected. Hence, despite vast genetic differences between the vaccine and both the challenge viruses, IN vaccination with NanoS100-SwIAV induced antigen-specific moderate levels of cross-protective immune responses.
Keywords: ADU-S100; Nano11; cell-mediated immune responses; intranasal vaccination; memory responses; swine; swine influenza A virus.
Conflict of interest statement
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures

Similar articles
-
A split influenza vaccine formulated with a combination adjuvant composed of alpha-D-glucan nanoparticles and a STING agonist elicits cross-protective immunity in pigs.J Nanobiotechnology. 2022 Nov 11;20(1):477. doi: 10.1186/s12951-022-01677-2. J Nanobiotechnology. 2022. PMID: 36369044 Free PMC article.
-
Intranasal Delivery of Inactivated Influenza Virus and Poly(I:C) Adsorbed Corn-Based Nanoparticle Vaccine Elicited Robust Antigen-Specific Cell-Mediated Immune Responses in Maternal Antibody Positive Nursery Pigs.Front Immunol. 2020 Dec 16;11:596964. doi: 10.3389/fimmu.2020.596964. eCollection 2020. Front Immunol. 2020. PMID: 33391267 Free PMC article.
-
Immunity and Protective Efficacy of Mannose Conjugated Chitosan-Based Influenza Nanovaccine in Maternal Antibody Positive Pigs.Front Immunol. 2021 Mar 4;12:584299. doi: 10.3389/fimmu.2021.584299. eCollection 2021. Front Immunol. 2021. PMID: 33746943 Free PMC article.
-
Characterization of a novel functional porcine CD3+CD4lowCD8α+CD8β+ T-helper/memory lymphocyte subset in the respiratory tract lymphoid tissues of swine influenza A virus vaccinated pigs.Vet Immunol Immunopathol. 2024 Aug;274:110785. doi: 10.1016/j.vetimm.2024.110785. Epub 2024 Jun 4. Vet Immunol Immunopathol. 2024. PMID: 38861830
-
Are pigs overestimated as a source of zoonotic influenza viruses?Porcine Health Manag. 2022 Jun 30;8(1):30. doi: 10.1186/s40813-022-00274-x. Porcine Health Manag. 2022. PMID: 35773676 Free PMC article. Review.
References
-
- Hayward A.C., Wang L., Goonetilleke N., Fragaszy E.B., Bermingham A., Copas A., Dukes O., Millett E.R., Nazareth I., Nguyen-Van-Tam J.S., et al. Natural T Cell-mediated Protection against Seasonal and Pandemic Influenza. Results of the Flu Watch Cohort Study. Am. J. Respir. Crit. Care Med. 2015;191:1422–1431. doi: 10.1164/rccm.201411-1988OC. - DOI - PMC - PubMed
-
- Gras S., Kedzierski L., Valkenburg S.A., Laurie K., Liu Y.C., Denholm J.T., Richards M.J., Rimmelzwaan G.F., Kelso A., Doherty P.C., et al. Cross-reactive CD8+ T-cell immunity between the pandemic H1N1-2009 and H1N1-1918 influenza A viruses. Proc. Natl. Acad. Sci. USA. 2010;107:12599–12604. doi: 10.1073/pnas.1007270107. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
