

Ancient diversity in host-parasite interaction genes in a model parasitic nematode
- PMID: 38012132
- PMCID: PMC10682056
- DOI: 10.1038/s41467-023-43556-w
Ancient diversity in host-parasite interaction genes in a model parasitic nematode
Abstract
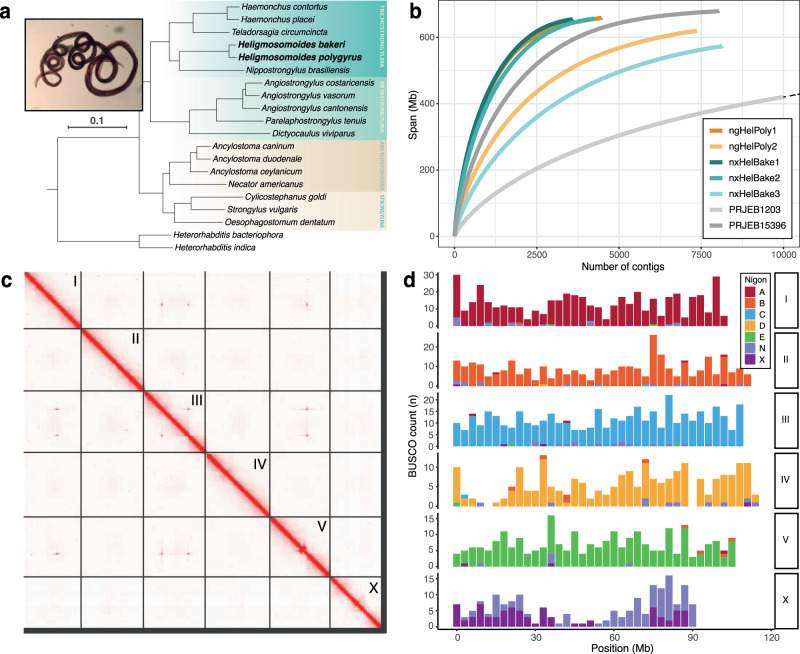
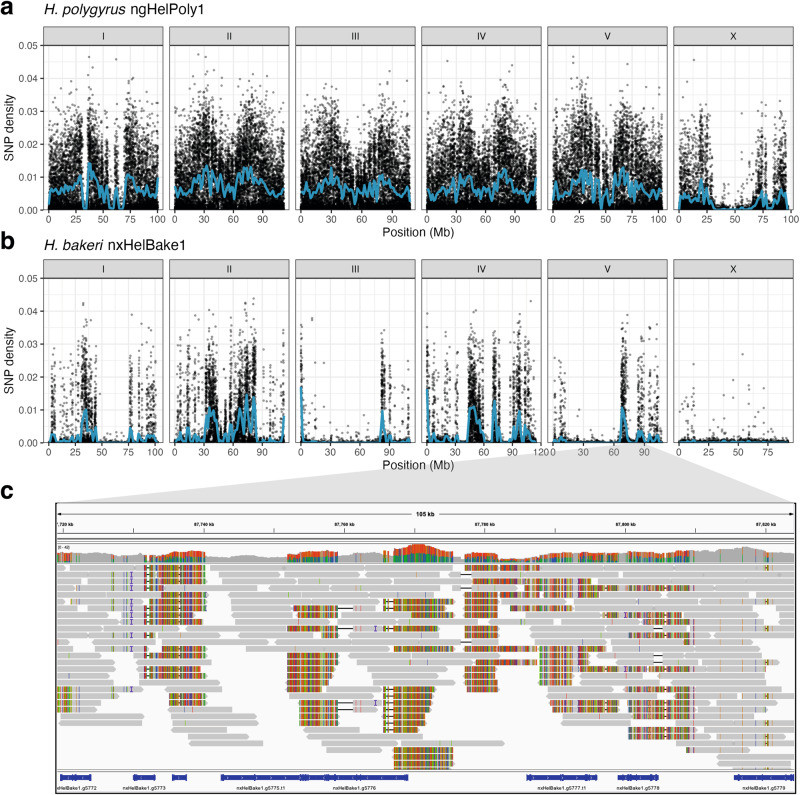
Host-parasite interactions exert strong selection pressures on the genomes of both host and parasite. These interactions can lead to negative frequency-dependent selection, a form of balancing selection that is hypothesised to explain the high levels of polymorphism seen in many host immune and parasite antigen loci. Here, we sequence the genomes of several individuals of Heligmosomoides bakeri, a model parasite of house mice, and Heligmosomoides polygyrus, a closely related parasite of wood mice. Although H. bakeri is commonly referred to as H. polygyrus in the literature, their genomes show levels of divergence that are consistent with at least a million years of independent evolution. The genomes of both species contain hyper-divergent haplotypes that are enriched for proteins that interact with the host immune response. Many of these haplotypes originated prior to the divergence between H. bakeri and H. polygyrus, suggesting that they have been maintained by long-term balancing selection. Together, our results suggest that the selection pressures exerted by the host immune response have played a key role in shaping patterns of genetic diversity in the genomes of parasitic nematodes.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Heligmosomoides bakeri: a new name for an old worm?Trends Parasitol. 2010 Nov;26(11):524-9. doi: 10.1016/j.pt.2010.07.001. Epub 2010 Aug 21. Trends Parasitol. 2010. PMID: 20729145
-
Molecular evidence that Heligmosomoides polygyrus from laboratory mice and wood mice are separate species.Parasitology. 2006 Jul;133(Pt 1):111-22. doi: 10.1017/S0031182006000047. Epub 2006 Mar 15. Parasitology. 2006. PMID: 16536883
-
Experience-dependent olfactory behaviors of the parasitic nematode Heligmosomoides polygyrus.PLoS Pathog. 2017 Nov 30;13(11):e1006709. doi: 10.1371/journal.ppat.1006709. eCollection 2017 Nov. PLoS Pathog. 2017. PMID: 29190282 Free PMC article.
-
Understanding the role of antibodies in murine infections with Heligmosomoides (polygyrus) bakeri: 35 years ago, now and 35 years ahead.Parasite Immunol. 2014 Mar;36(3):115-24. doi: 10.1111/pim.12057. Parasite Immunol. 2014. PMID: 23889357 Review.
-
Heligmosomoides bakeri: a model for exploring the biology and genetics of resistance to chronic gastrointestinal nematode infections.Parasitology. 2009 Oct;136(12):1565-80. doi: 10.1017/S0031182009006003. Epub 2009 May 19. Parasitology. 2009. PMID: 19450375 Review.
Cited by
-
Multiple Displacement Amplification Facilitates SMRT Sequencing of Microscopic Animals and the Genome of the Gastrotrich Lepidodermella squamata (Dujardin 1841).Genome Biol Evol. 2024 Dec 4;16(12):evae254. doi: 10.1093/gbe/evae254. Genome Biol Evol. 2024. PMID: 39590608 Free PMC article.
-
Quantitative assessment of spicule length in Heligmosomoides spp. (Nematoda, Heligmosomidae): distinction between H. bakeri, H. polygyrus and H. glareoli.Parasitology. 2023 Sep;150(11):1022-1030. doi: 10.1017/S0031182023000872. Epub 2023 Sep 14. Parasitology. 2023. PMID: 37705292 Free PMC article.
-
Genomic analysis of the zooplankton-associated pathogenic bacterium Spirobacillus cienkowskii reveals its functional and metabolic capacities.Microb Genom. 2025 Aug;11(8):001463. doi: 10.1099/mgen.0.001463. Microb Genom. 2025. PMID: 40758756 Free PMC article.
-
Hookworm genes encoding intestinal excreted-secreted proteins are transcriptionally upregulated in response to the host's immune system.bioRxiv [Preprint]. 2025 Feb 3:2025.02.01.636063. doi: 10.1101/2025.02.01.636063. bioRxiv. 2025. PMID: 39975173 Free PMC article. Preprint.
-
Heligmosomoides bakeri and Toxoplasma gondii co-infection leads to increased mortality associated with changes in immune resistance in the lymphoid compartment and disease pathology.PLoS One. 2024 Jul 1;19(7):e0292408. doi: 10.1371/journal.pone.0292408. eCollection 2024. PLoS One. 2024. PMID: 38950025 Free PMC article.
References
-
- Van Valen L. A new evolutionary law. Evol. Theory. 1973;1:1–30.
-
- Tellier A, Moreno‐Gámez S, Stephan W. Speed Of Adaptation And Genomic Footprints Of Host–Parasite Coevolution Under Arms Race And Trench Warfare Dynamics. Evolution. 2014;68:2211–2224. - PubMed
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
