

Genetically encoding multiple functionalities into extracellular vesicles for the targeted delivery of biologics to T cells
- PMID: 38012307
- PMCID: PMC11088532
- DOI: 10.1038/s41551-023-01142-x
Genetically encoding multiple functionalities into extracellular vesicles for the targeted delivery of biologics to T cells
Abstract
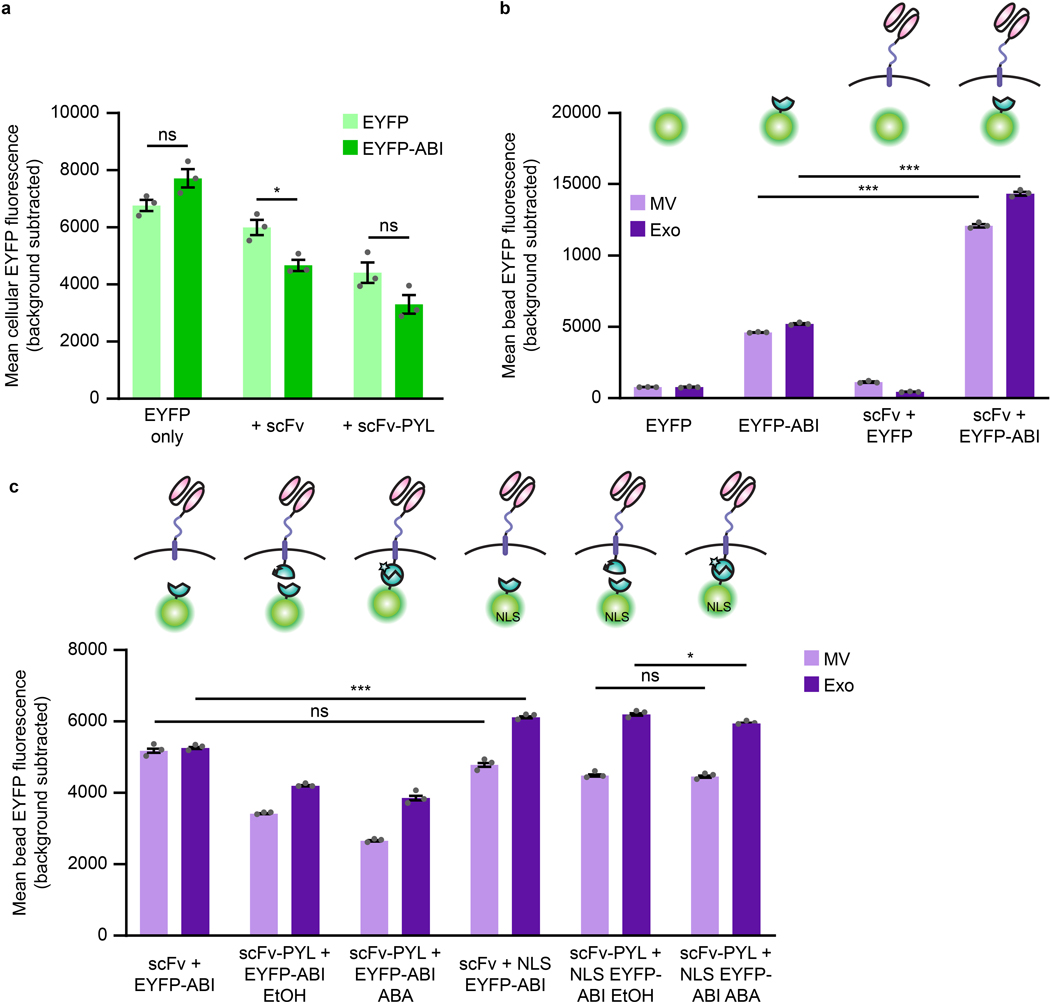
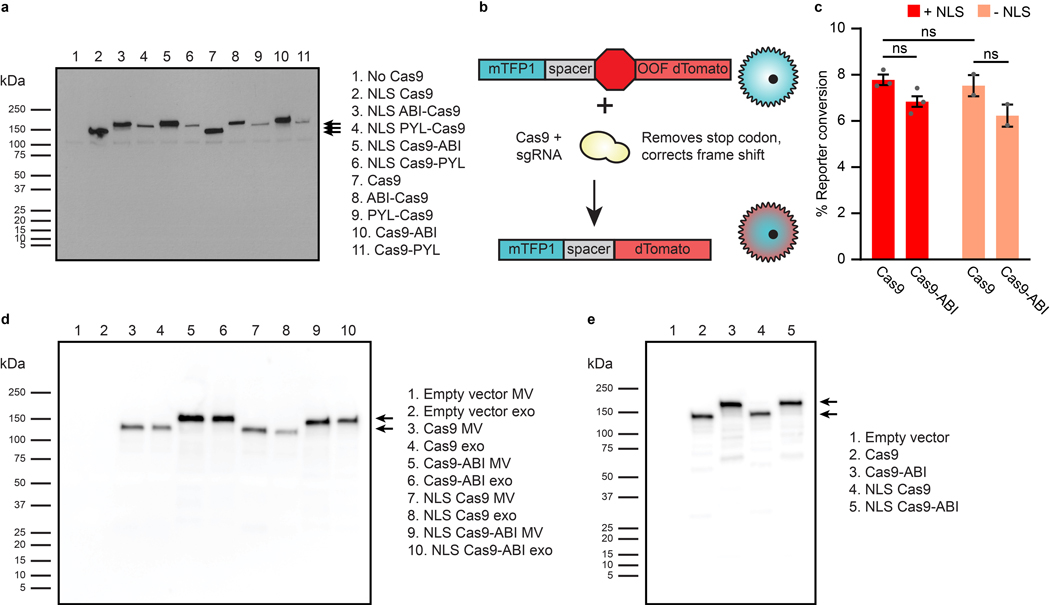
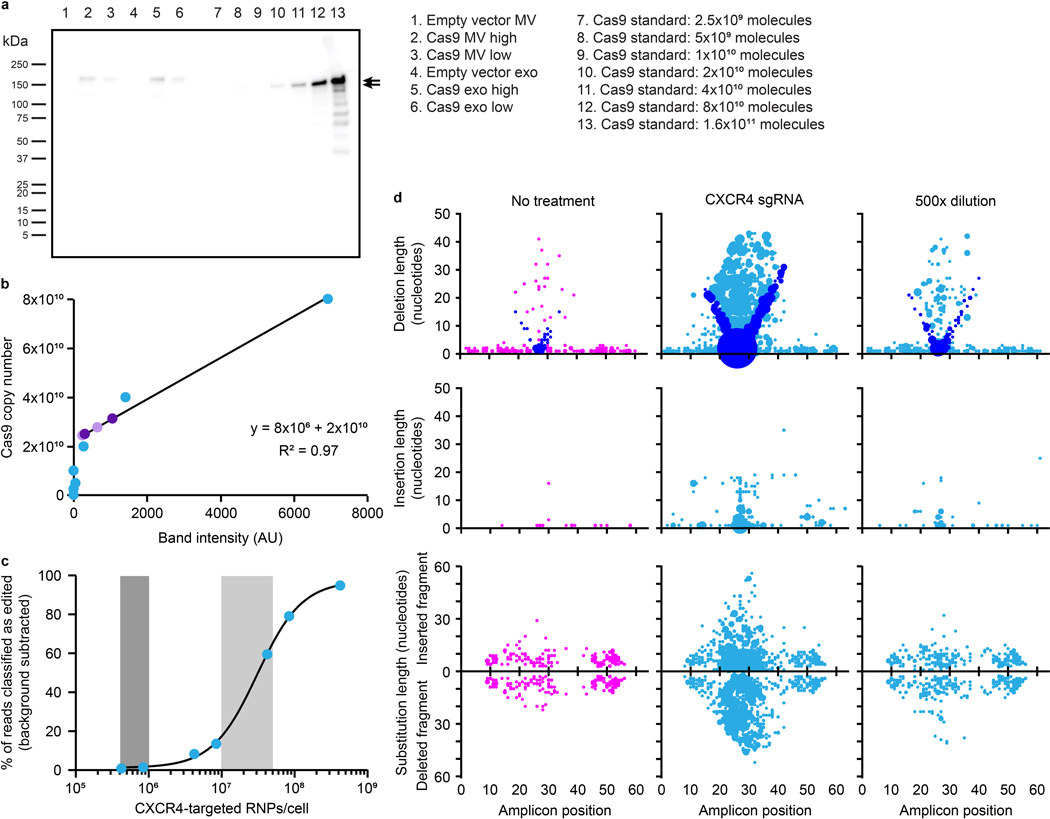
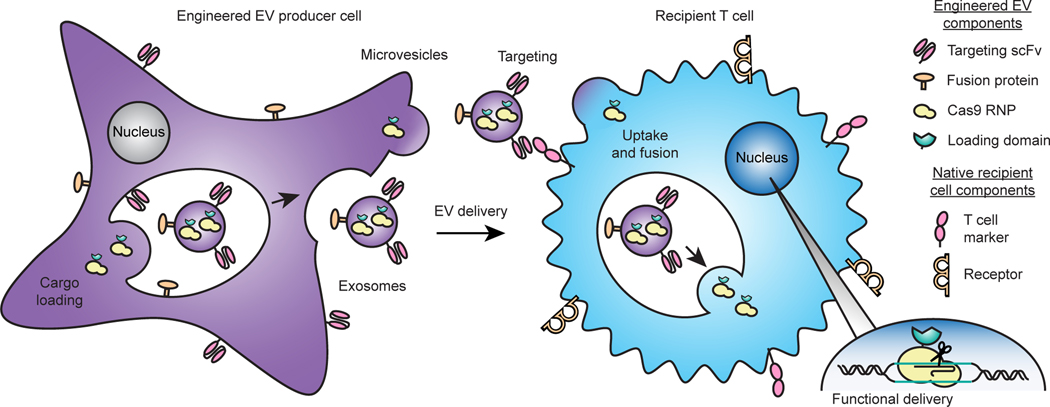
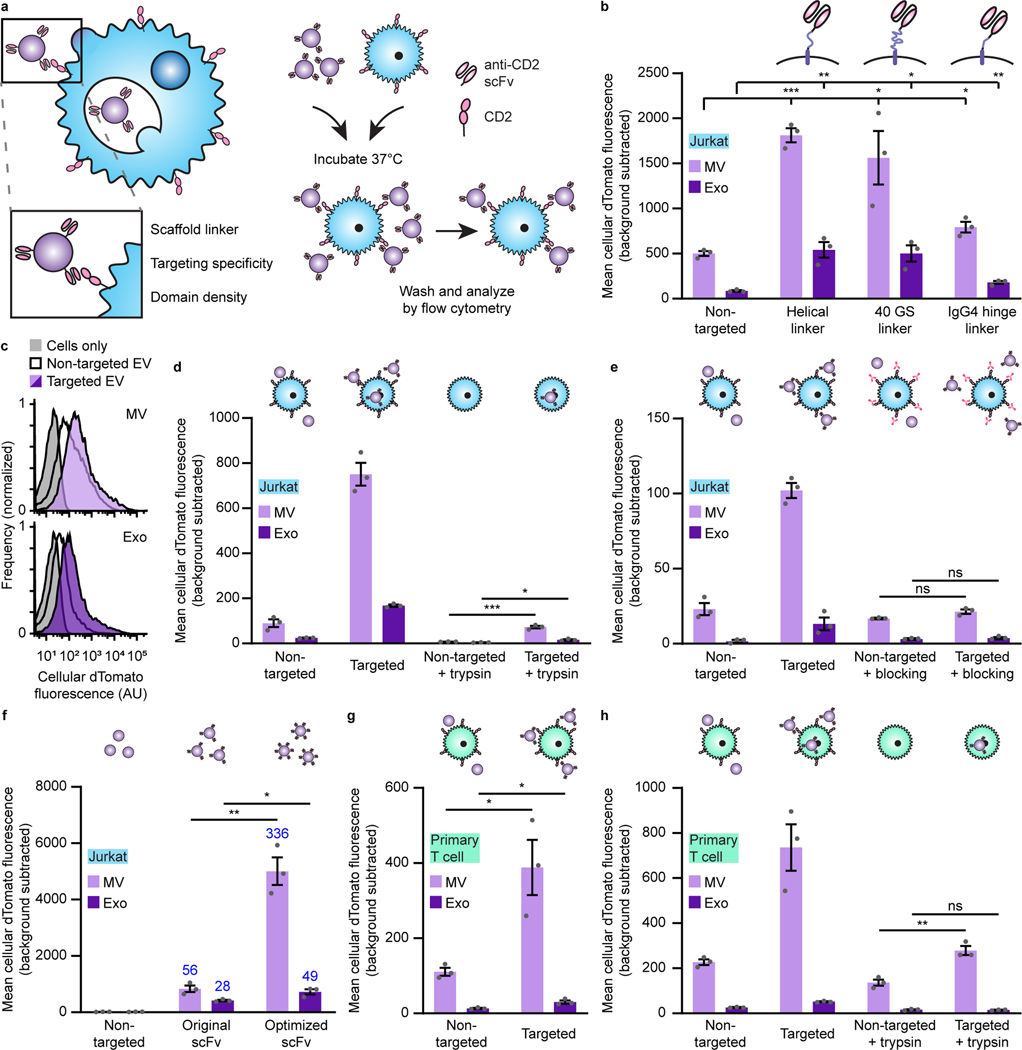
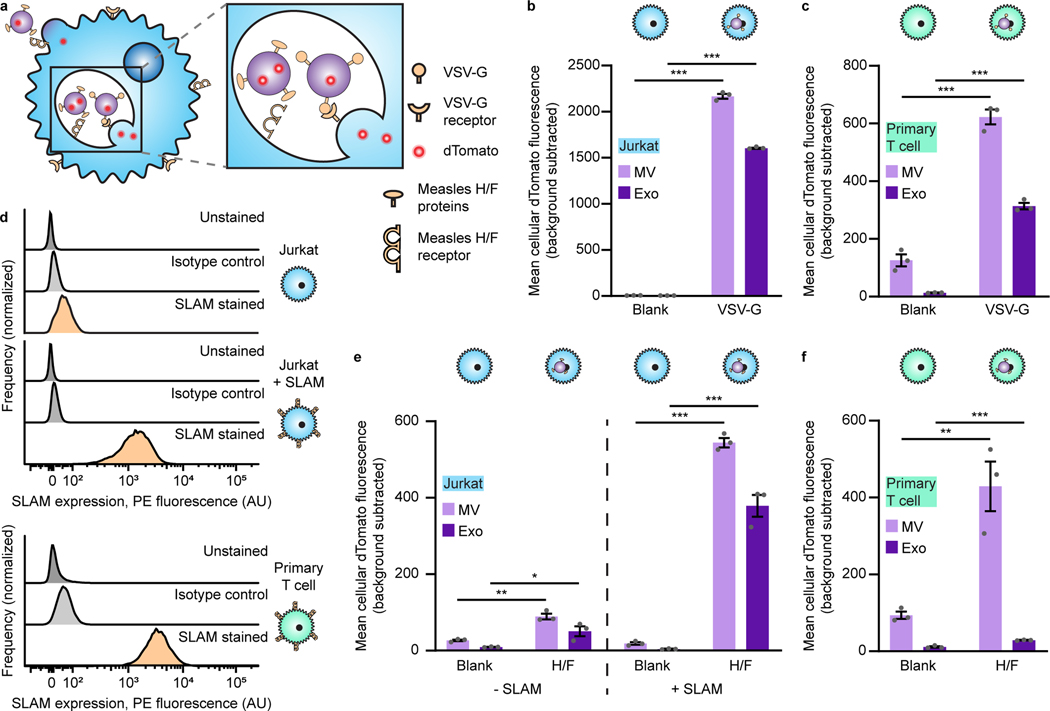
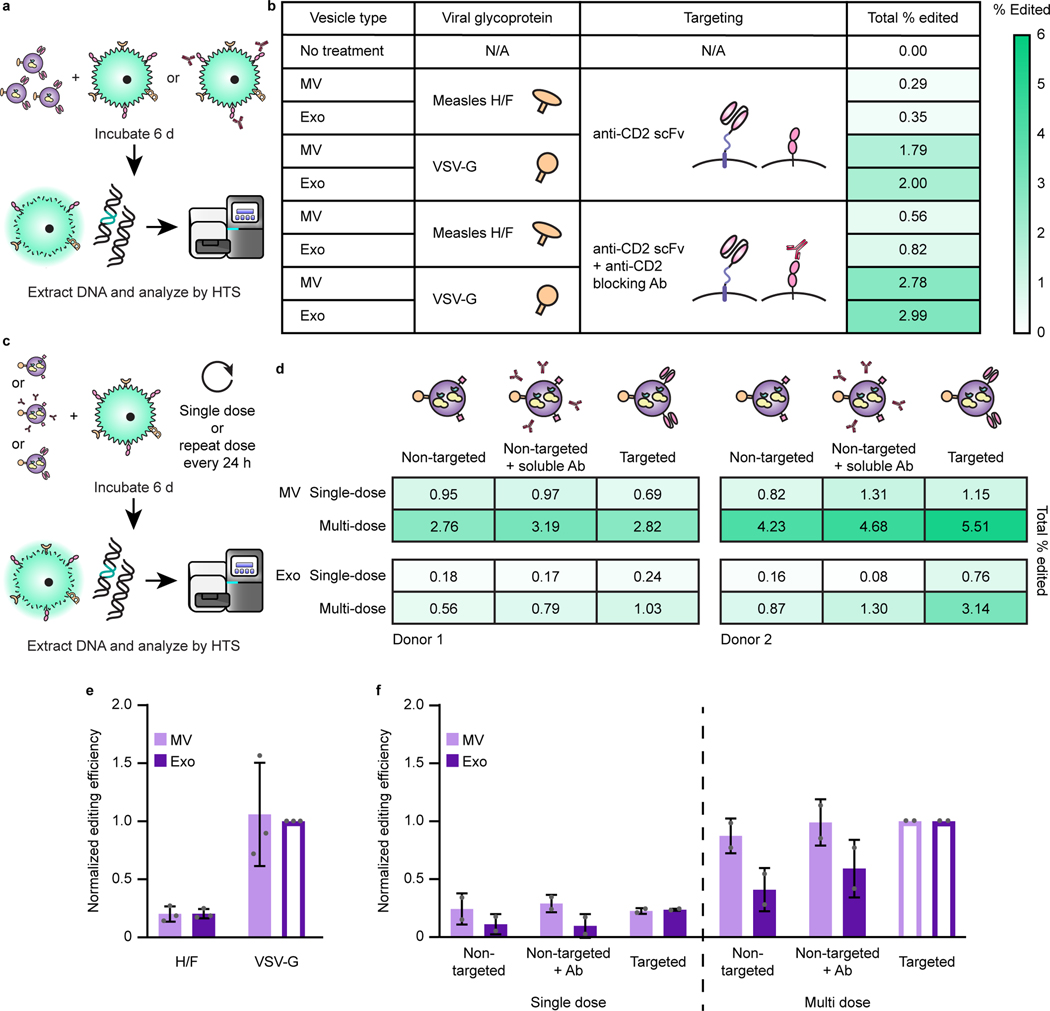
The genetic modification of T cells has advanced cellular immunotherapies, yet the delivery of biologics specifically to T cells remains challenging. Here we report a suite of methods for the genetic engineering of cells to produce extracellular vesicles (EVs)-which naturally encapsulate and transfer proteins and nucleic acids between cells-for the targeted delivery of biologics to T cells without the need for chemical modifications. Specifically, the engineered cells secreted EVs that actively loaded protein cargo via a protein tag and that displayed high-affinity T-cell-targeting domains and fusogenic glycoproteins. We validated the methods by engineering EVs that delivered Cas9-single-guide-RNA complexes to ablate the gene encoding the C-X-C chemokine co-receptor type 4 in primary human CD4+ T cells. The strategy is amenable to the targeted delivery of biologics to other cell types.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
J.N.L. and D.M.S. are co-inventors on patent pending intellectual property that covers some technologies reported in this manuscript. J.N.L. and D.M.S. have financial interest in Syenex, which could potentially benefit from the outcomes of this research.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
