

This is a preprint.
Single phage proteins sequester TIR- and cGAS-generated signaling molecules
- PMID: 38014003
- PMCID: PMC10680739
- DOI: 10.1101/2023.11.15.567273
Single phage proteins sequester TIR- and cGAS-generated signaling molecules
Update in
-
Single phage proteins sequester signals from TIR and cGAS-like enzymes.Nature. 2024 Nov;635(8039):719-727. doi: 10.1038/s41586-024-08122-4. Epub 2024 Oct 30. Nature. 2024. PMID: 39478223 Free PMC article.
Abstract
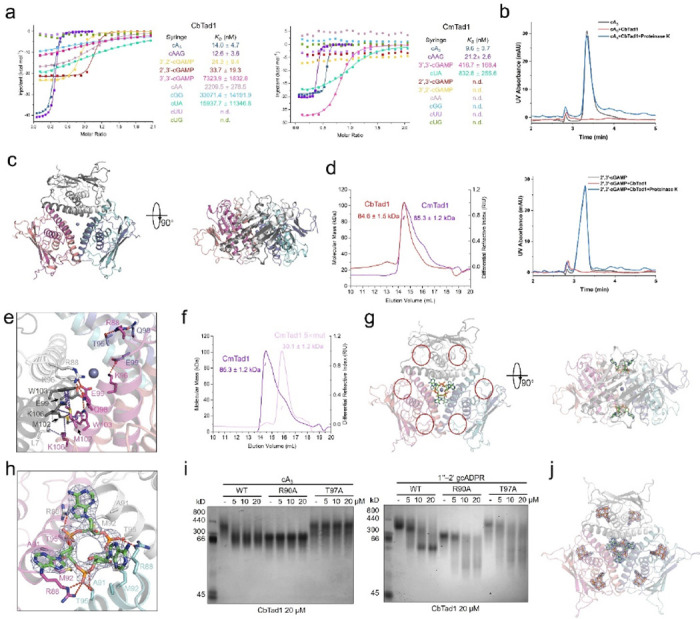
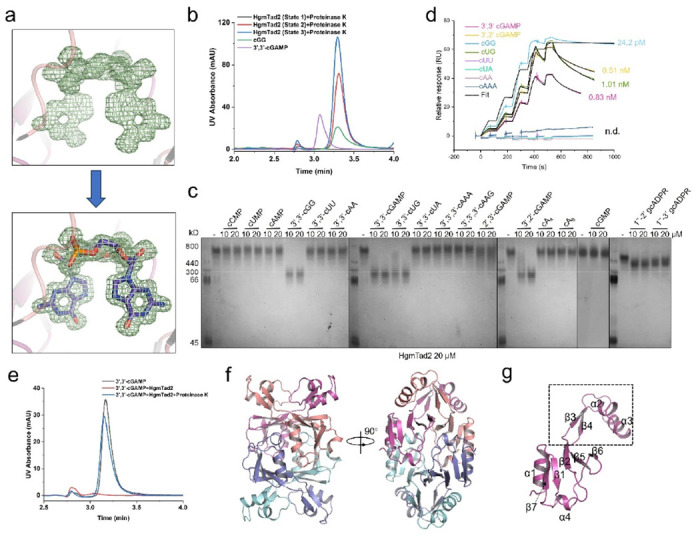
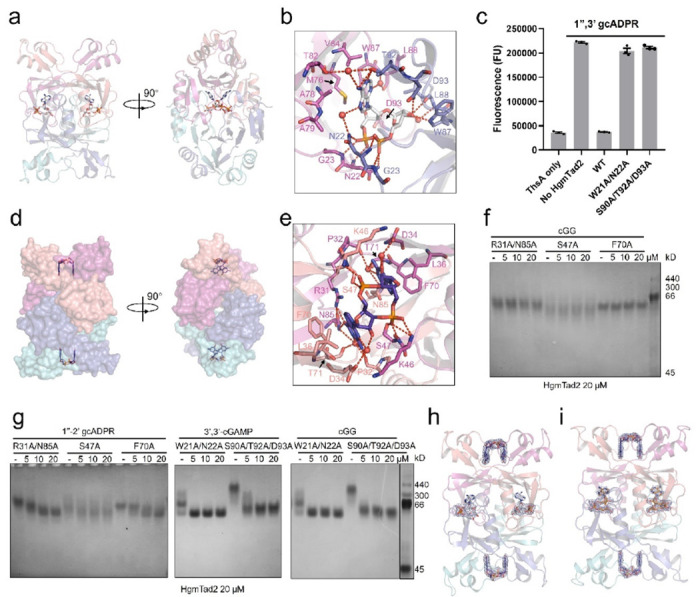
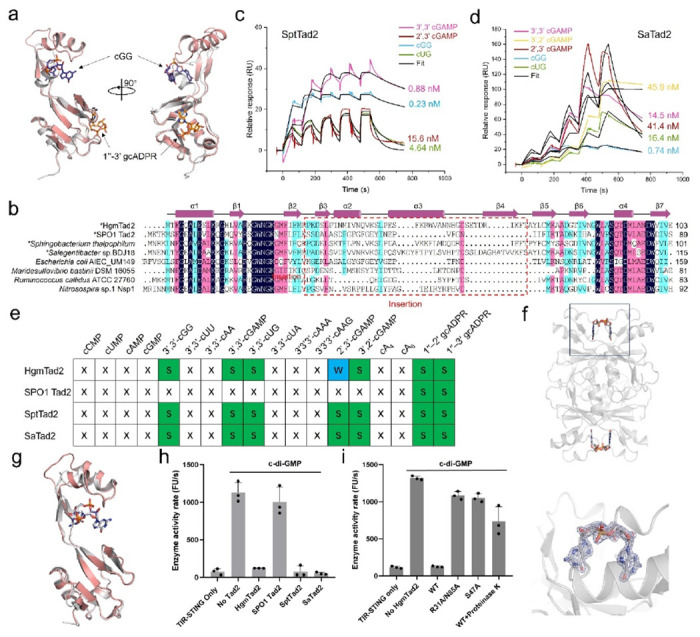
Prokaryotic anti-phage immune systems use TIR (toll/interleukin-1 receptor) and cGAS (cyclic GMP-AMP synthase) enzymes to produce 1"-3'/1"-2' glycocyclic ADPR (gcADPR) and cyclid di-/trinucleotides (CDNs and CTNs) signaling molecules that limit phage replication, respectively 1-3. However, how phages neutralize these common systems is largely unknown. Here, we show that Thoeris anti-defense proteins Tad1 4 and Tad2 5 both have anti-CBASS activity by simultaneously sequestering CBASS cyclic oligonucleotides. Strikingly, apart from binding Thoeris signals 1"-3' and 1"-2' gcADPR, Tad1 also binds numerous CBASS CDNs/CTNs with high affinity, inhibiting CBASS systems using these molecules in vivo and in vitro. The hexameric Tad1 has six binding sites for CDNs or gcADPR, which are independent from two high affinity binding sites for CTNs. Tad2 also sequesters various CDNs in addition to gcADPR molecules, inhibiting CBASS systems using these CDNs. However, the binding pockets for CDNs and gcADPR are different in Tad2, whereby a tetramer can bind two CDNs and two gcADPR molecules simultaneously. Taken together, Tad1 and Tad2 are both two-pronged inhibitors that, alongside anti-CBASS protein 2, establish a paradigm of phage proteins that flexibly sequester a remarkable breadth of cyclic nucleotides involved in TIR- and cGAS-based anti-phage immunity.
Conflict of interest statement
J.B.-D. is a scientific advisory board member of SNIPR Biome and Excision Biotherapeutics, a consultant to LeapFrog Bio and BiomX, and a scientific advisory board member and co-founder of Acrigen Biosciences. The Bondy-Denomy lab received research support from Felix Biotechnology.
Figures


Similar articles
-
Single phage proteins sequester signals from TIR and cGAS-like enzymes.Nature. 2024 Nov;635(8039):719-727. doi: 10.1038/s41586-024-08122-4. Epub 2024 Oct 30. Nature. 2024. PMID: 39478223 Free PMC article.
-
Phage anti-CBASS protein simultaneously sequesters cyclic trinucleotides and dinucleotides.Mol Cell. 2024 Jan 18;84(2):375-385.e7. doi: 10.1016/j.molcel.2023.11.026. Epub 2023 Dec 15. Mol Cell. 2024. PMID: 38103556 Free PMC article.
-
Viruses inhibit TIR gcADPR signalling to overcome bacterial defence.Nature. 2022 Nov;611(7935):326-331. doi: 10.1038/s41586-022-05375-9. Epub 2022 Sep 29. Nature. 2022. PMID: 36174646
-
Sequestering survival: sponge-like proteins in phage evasion of bacterial immune defenses.Front Immunol. 2025 Apr 17;16:1545308. doi: 10.3389/fimmu.2025.1545308. eCollection 2025. Front Immunol. 2025. PMID: 40313938 Free PMC article. Review.
-
The arms race between bacteria CBASS and bacteriophages.Front Immunol. 2023 Jul 28;14:1224341. doi: 10.3389/fimmu.2023.1224341. eCollection 2023. Front Immunol. 2023. PMID: 37575224 Free PMC article. Review.
References
-
- Manik M.K. et al. Cyclic ADP ribose isomers: Production, chemical structures, and immune signaling. Science 377, eadc8969 (2022). - PubMed
-
- Ofir G. et al. Antiviral activity of bacterial TIR domains via immune signalling molecules. Nature 600, 116–120 (2021). - PubMed
-
- Cohen D. et al. Cyclic GMP-AMP signalling protects bacteria against viral infection. Nature 574, 691–695 (2019). - PubMed
-
- Leavitt A. et al. Viruses inhibit TIR gcADPR signaling to overcome bacterial defense. Nature 611, 326–331 (2022). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources