

This is a preprint.
The Dmc1 recombinase physically interacts with and promotes the meiotic crossover functions of the Mlh1-Mlh3 endonuclease
- PMID: 38014100
- PMCID: PMC10680668
- DOI: 10.1101/2023.11.13.566911
The Dmc1 recombinase physically interacts with and promotes the meiotic crossover functions of the Mlh1-Mlh3 endonuclease
Update in
-
The Dmc1 recombinase physically interacts with and promotes the meiotic crossover functions of the Mlh1-Mlh3 endonuclease.Genetics. 2024 Jul 8;227(3):iyae066. doi: 10.1093/genetics/iyae066. Genetics. 2024. PMID: 38657110 Free PMC article.
Abstract
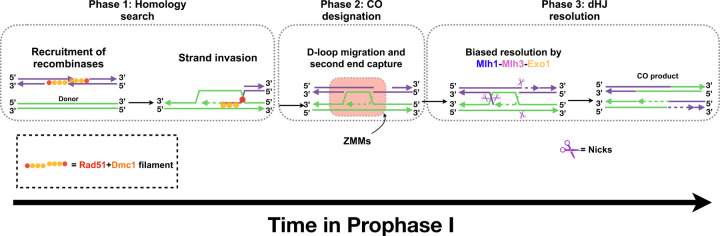
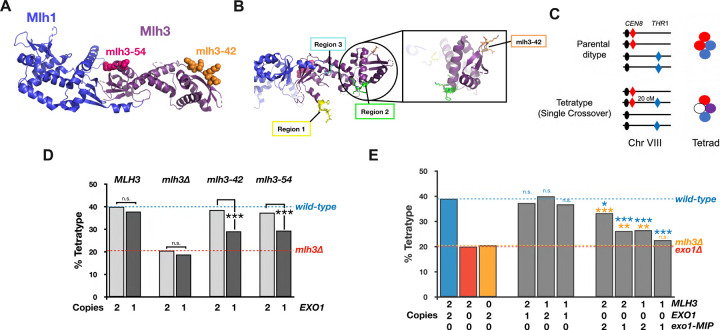
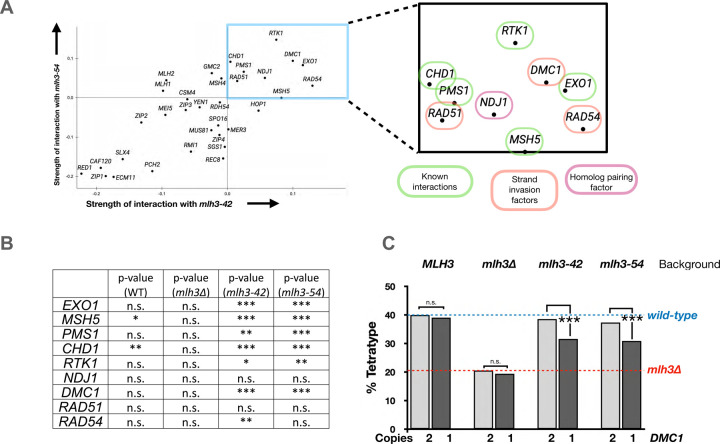
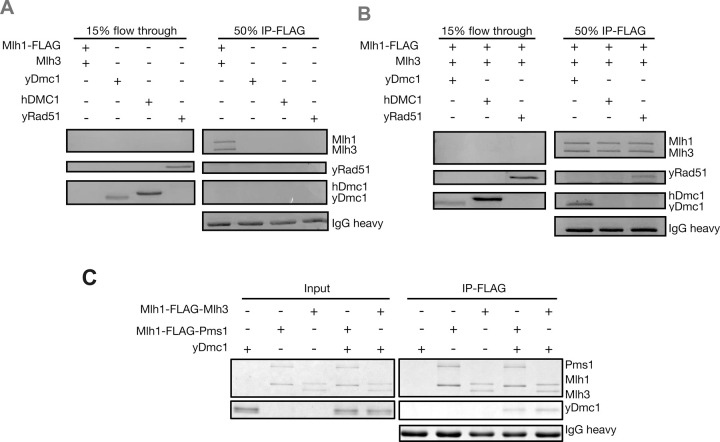
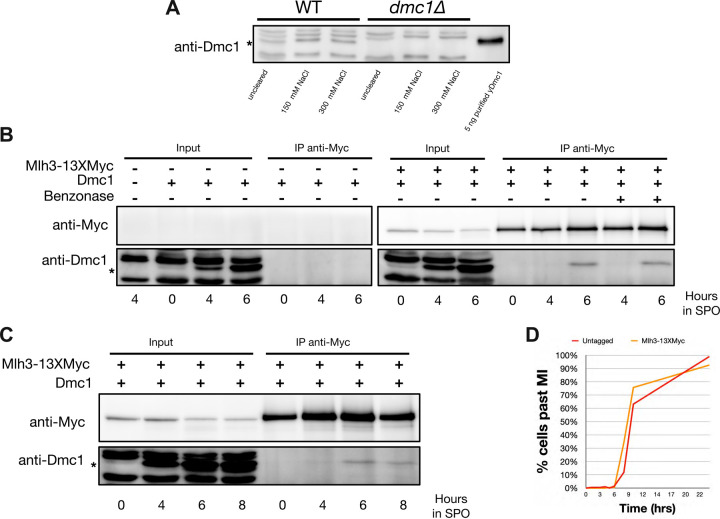
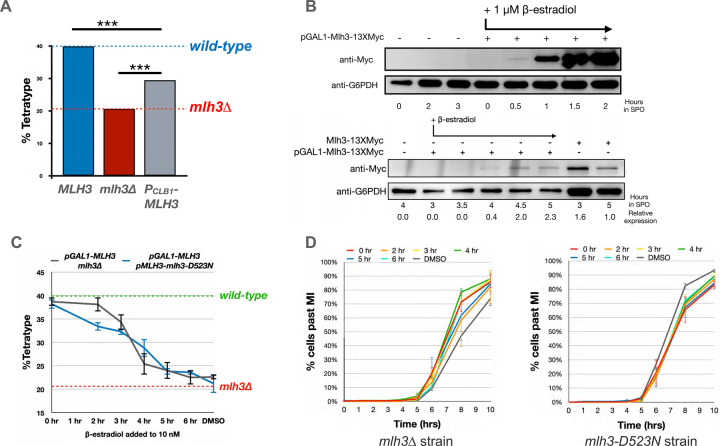
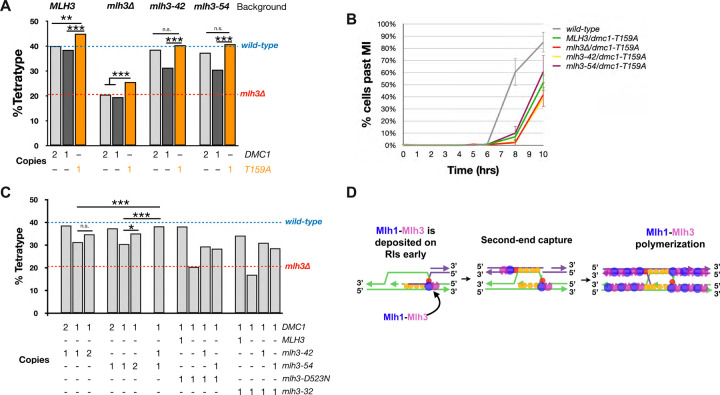
The accurate segregation of homologous chromosomes during the Meiosis I reductional division in most sexually reproducing eukaryotes requires crossing over between homologs. In baker's yeast approximately 80 percent of meiotic crossovers result from Mlh1-Mlh3 and Exo1 acting to resolve double-Holliday junction (dHJ) intermediates in a biased manner. Little is known about how Mlh1-Mlh3 is recruited to recombination intermediates and whether it interacts with other meiotic factors prior to its role in crossover resolution. We performed a haploinsufficiency screen in baker's yeast to identify novel genetic interactors with Mlh1-Mlh3 using sensitized mlh3 alleles that disrupt the stability of the Mlh1-Mlh3 complex and confer defects in mismatch repair but do not disrupt meiotic crossing over. We identified several genetic interactions between MLH3 and DMC1, the recombinase responsible for recombination between homologous chromosomes during meiosis. We then showed that Mlh3 physically interacts with Dmc1 in vitro and at times in meiotic prophase when Dmc1 acts as a recombinase. Interestingly, restricting MLH3 expression to roughly the time of crossover resolution resulted in a mlh3 null-like phenotype for crossing over. Our data are consistent with a model in which Dmc1 nucleates a polymer of Mlh1-Mlh3 to promote crossing over.
Keywords: Dmc1; Holliday junction resolution; Mlh1; Mlh3; crossing over; meiotic recombination.
Conflict of interest statement
Conflicts of interest. None declared.
Figures
References
-
- Abdullah MF, Hoffmann ER, Cotton VE, Borts RH. 2004. A role for the MutL homologue MLH2 in controlling heteroduplex formation and in regulating between two different crossover pathways in budding yeast. Cytogenet Genome Res. 107:180–190. - PubMed
-
- Allers T, Lichten M. 2001. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell. 106:47–57. - PubMed
-
- Altmannova V, Firlej M, Müller F, Janning P, Rauleder R, Rousova D, Schäffler A, Bange T, Weir JR. 2023. Biochemical characterisation of Mer3 helicase interactions and the protection of meiotic recombination intermediates. Nucleic Acids Res. 51:4363–4384. doi: 10.1093/nar/gkad175. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources