

This is a preprint.
Creation of de novo cryptic splicing for ALS/FTD precision medicine
- PMID: 38014203
- PMCID: PMC10680699
- DOI: 10.1101/2023.11.15.565967
Creation of de novo cryptic splicing for ALS/FTD precision medicine
Update in
-
Creation of de novo cryptic splicing for ALS and FTD precision medicine.Science. 2024 Oct 4;386(6717):61-69. doi: 10.1126/science.adk2539. Epub 2024 Oct 3. Science. 2024. PMID: 39361759 Free PMC article.
Abstract
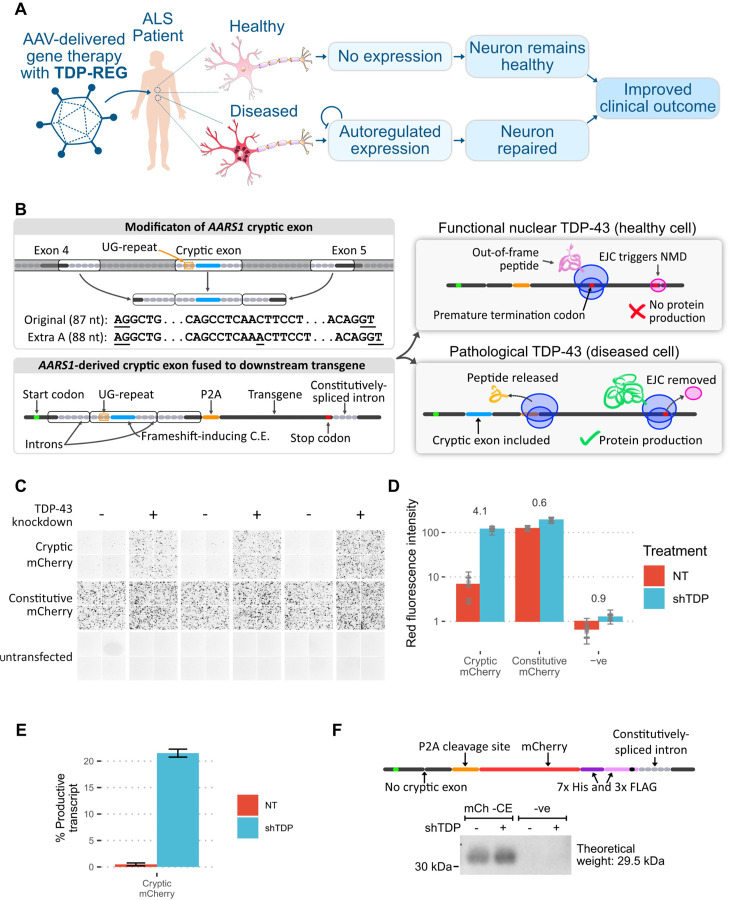
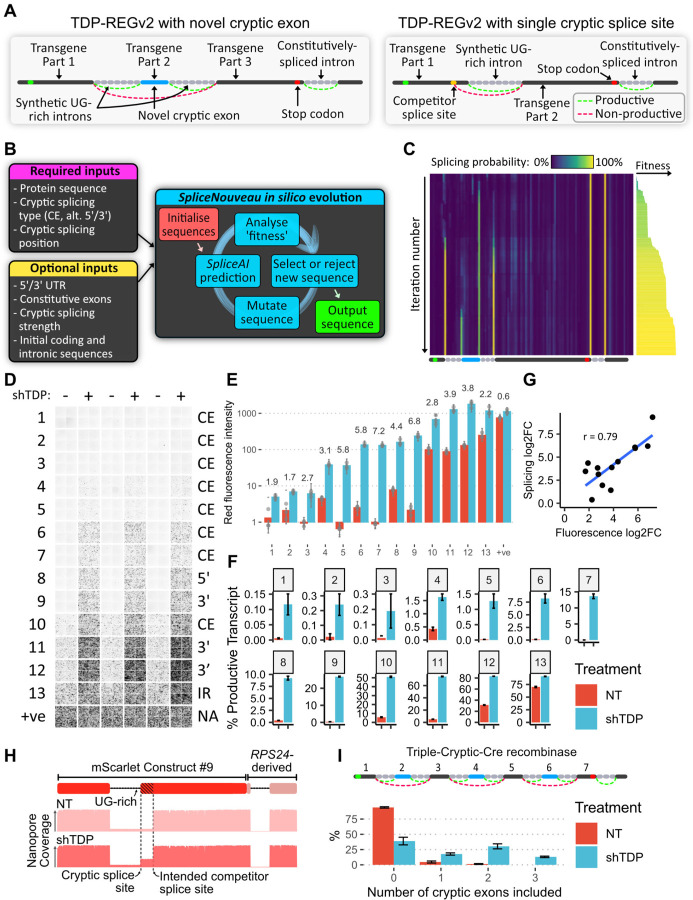
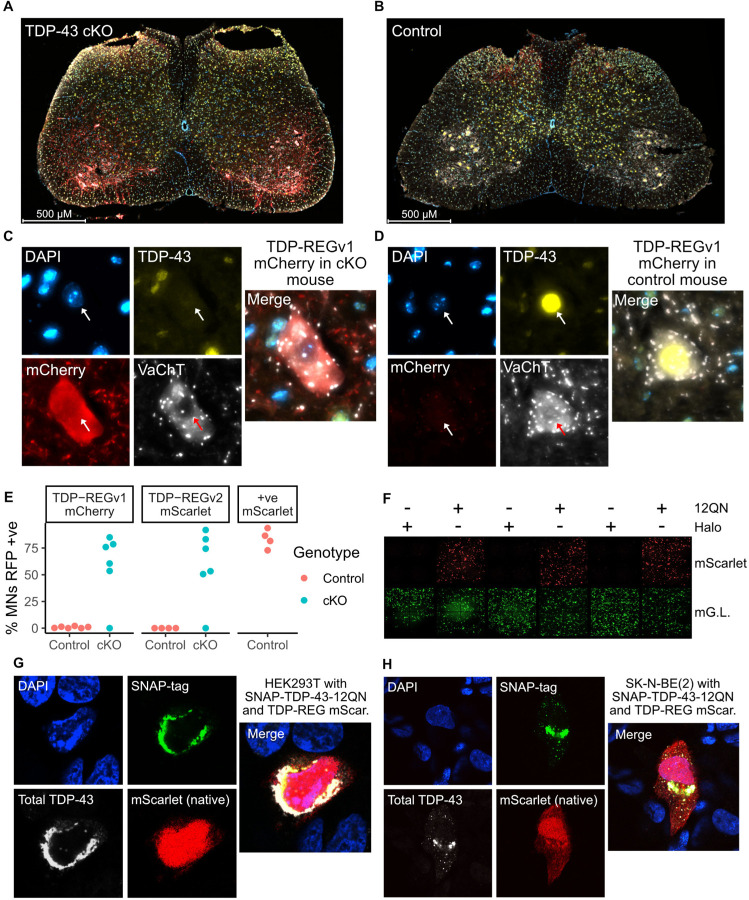
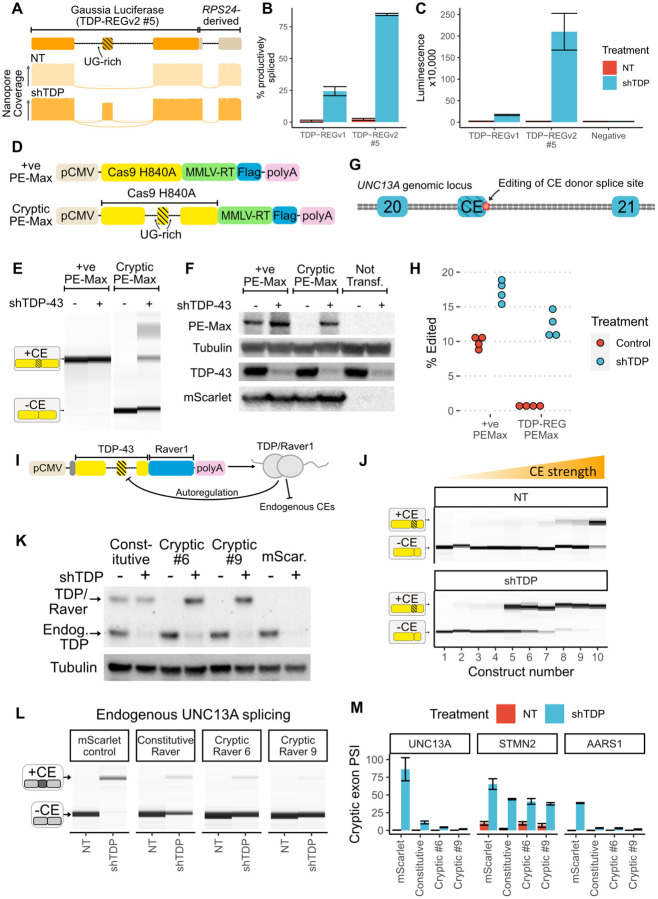
A system enabling the expression of therapeutic proteins specifically in diseased cells would be transformative, providing greatly increased safety and the possibility of pre-emptive treatment. Here we describe "TDP-REG", a precision medicine approach primarily for amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD), which exploits the cryptic splicing events that occur in cells with TDP-43 loss-of-function (TDP-LOF) in order to drive expression specifically in diseased cells. In addition to modifying existing cryptic exons for this purpose, we develop a deep-learning-powered algorithm for generating customisable cryptic splicing events, which can be embedded within virtually any coding sequence. By placing part of a coding sequence within a novel cryptic exon, we tightly couple protein expression to TDP-LOF. Protein expression is activated by TDP-LOF in vitro and in vivo, including TDP-LOF induced by cytoplasmic TDP-43 aggregation. In addition to generating a variety of fluorescent and luminescent reporters, we use this system to perform TDP-LOF-dependent genomic prime editing to ablate the UNC13A cryptic donor splice site. Furthermore, we design a panel of tightly gated, autoregulating vectors encoding a TDP-43/Raver1 fusion protein, which rescue key pathological cryptic splicing events. In summary, we combine deep-learning and rational design to create sophisticated splicing sensors, resulting in a platform that provides far safer therapeutics for neurodegeneration, potentially even enabling preemptive treatment of at-risk individuals.
Conflict of interest statement
Competing interests: PF and OGW have filed a patent application relating to TDP-REG technology. PF, ALB, MJK and OGW have filed a patent application relating to the use of ASO therapies for correcting cryptic splicing in UNC13A.
Figures
References
-
- Nelson P. T., Dickson D. W., Trojanowski J. Q., Jack C. R., Boyle P. A., Arfanakis K., Rademakers R., Alafuzoff I., Attems J., Brayne C., Coyle-Gilchrist I. T. S., Chui H. C., Fardo D. W., Flanagan M. E., Halliday G., Hokkanen S. R. K., Hunter S., Jicha G. A., Katsumata Y., Kawas C. H., Keene C. D., Kovacs G. G., Kukull W. A., Levey A. I., Makkinejad N., Montine T. J., Murayama S., Murray M. E., Nag S., Rissman R. A., Seeley W. W., Sperling R. A., White C. L. 3rd, Yu L., Schneider J. A., Limbic-predominant age-related TDP-43 encephalopathy (LATE): consensus working group report. Brain. 142, 1503–1527 (2019). - PMC - PubMed
-
- Klim J. R., Williams L. A., Limone F., Guerra San Juan I., Davis-Dusenbery B. N., Mordes D. A., Burberry A., Steinbaugh M. J., Gamage K. K., Kirchner R., Moccia R., Cassel S. H., Chen K., Wainger B. J., Woolf C. J., Eggan K., ALS-implicated protein TDP-43 sustains levels of STMN2, a mediator of motor neuron growth and repair. Nat. Neurosci. 22, 167–179 (2019). - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous