

This is a preprint.
Control of Biofilm Formation by an Agrobacterium tumefaciens Pterin-Binding Periplasmic Protein Conserved Among Pathogenic Bacteria
- PMID: 38014264
- PMCID: PMC10680838
- DOI: 10.1101/2023.11.18.567607
Control of Biofilm Formation by an Agrobacterium tumefaciens Pterin-Binding Periplasmic Protein Conserved Among Pathogenic Bacteria
Update in
-
Control of biofilm formation by an Agrobacterium tumefaciens pterin-binding periplasmic protein conserved among diverse Proteobacteria.Proc Natl Acad Sci U S A. 2024 Jun 18;121(25):e2319903121. doi: 10.1073/pnas.2319903121. Epub 2024 Jun 13. Proc Natl Acad Sci U S A. 2024. PMID: 38870058 Free PMC article.
Abstract
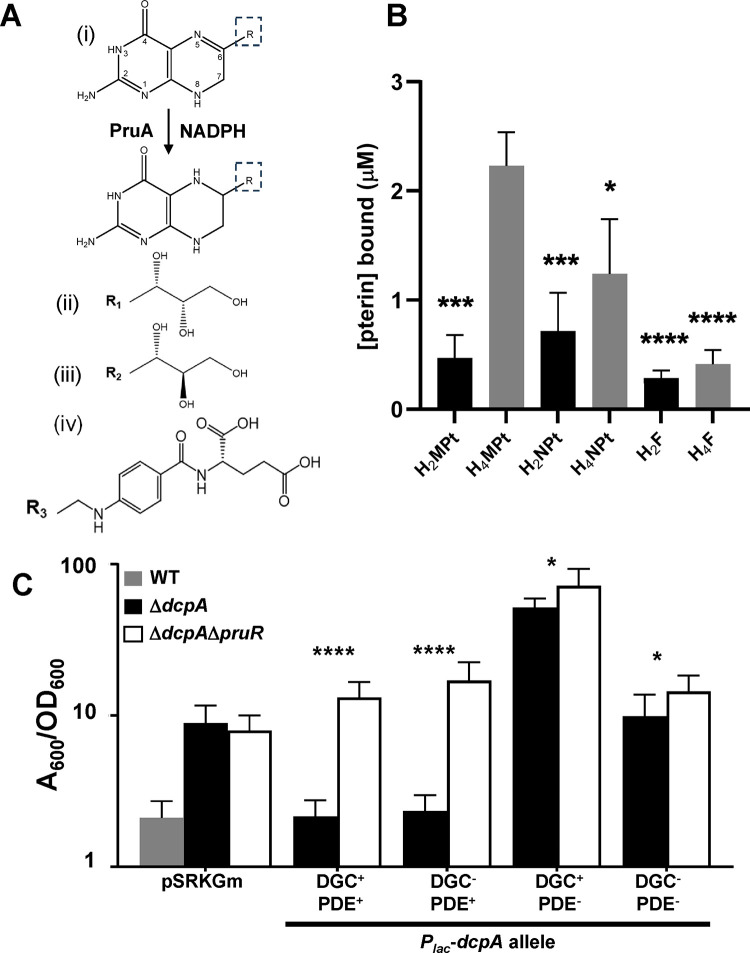
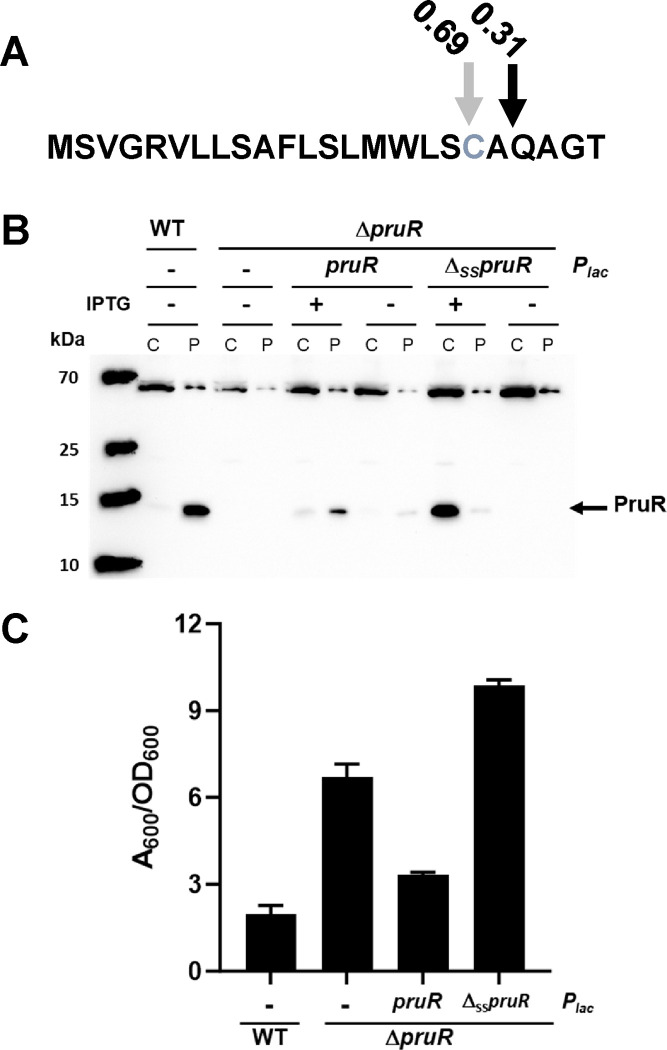
Biofilm formation and surface attachment in multiple Alphaproteobacteria is driven by unipolar polysaccharide (UPP) adhesins. The pathogen Agrobacterium tumefaciens produces a UPP adhesin, which is regulated by the intracellular second messenger cyclic diguanylate monophosphate (cdGMP). Prior studies revealed that DcpA, a diguanylate cyclase-phosphodiesterase (DGC-PDE), is crucial in control of UPP production and surface attachment. DcpA is regulated by PruR, a protein with distant similarity to enzymatic domains known to coordinate the molybdopterin cofactor (MoCo). Pterins are bicyclic nitrogen-rich compounds, several of which are formed via a non-essential branch of the folate biosynthesis pathway, distinct from MoCo. The pterin-binding protein PruR controls DcpA activity, fostering cdGMP breakdown and dampening its synthesis. Pterins are excreted and we report here that PruR associates with these metabolites in the periplasm, promoting interaction with the DcpA periplasmic domain. The pteridine reductase PruA, which reduces specific dihydro-pterin molecules to their tetrahydro forms, imparts control over DcpA activity through PruR. Tetrahydromonapterin preferentially associates with PruR relative to other related pterins, and the PruR-DcpA interaction is decreased in a pruA mutant. PruR and DcpA are encoded in an operon that is conserved amongst multiple Proteobacteria including mammalian pathogens. Crystal structures reveal that PruR and several orthologs adopt a conserved fold, with a pterin-specific binding cleft that coordinates the bicyclic pterin ring. These findings define a new pterin-responsive regulatory mechanism that controls biofilm formation and related cdGMP-dependent phenotypes in A. tumefaciens and is found in multiple additional bacterial pathogens.
Keywords: Biofilm; Biological Sciences; Microbiology; cyclic diguanylate monophosphate; protein structure; pterin; regulation.
Conflict of interest statement
Competing Interest Statement: K.J.F.S. has a significant interest in Situ Biosciences, a contract research organization that conducts research unrelated to this study. All other authors declare no conflicts of interest. The funders had no role in study design, data collection, and analysis, decision to publish, or preparation of the manuscript.
Figures





References
-
- Ciofu O., Moser C., Jensen P. O., Hoiby N., Tolerance and resistance of microbial biofilms. Nat Rev Microbiol 20, 621–635 (2022). - PubMed
-
- Berne C., Ellison C. K., Ducret A., Brun Y. V., Bacterial adhesion at the single-cell level. Nat Rev Microbiol 16, 616–627 (2018). - PubMed
-
- Jenal U., Reinders A., Lori C., Cyclic di-GMP: second messenger extraordinaire. Nat Rev Microbiol 15, 271–284 (2017). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources