

DYRK-family kinases regulate Candida albicans morphogenesis and virulence through the Ras1/PKA pathway
- PMID: 38015416
- PMCID: PMC10746247
- DOI: 10.1128/mbio.02183-23
DYRK-family kinases regulate Candida albicans morphogenesis and virulence through the Ras1/PKA pathway
Abstract
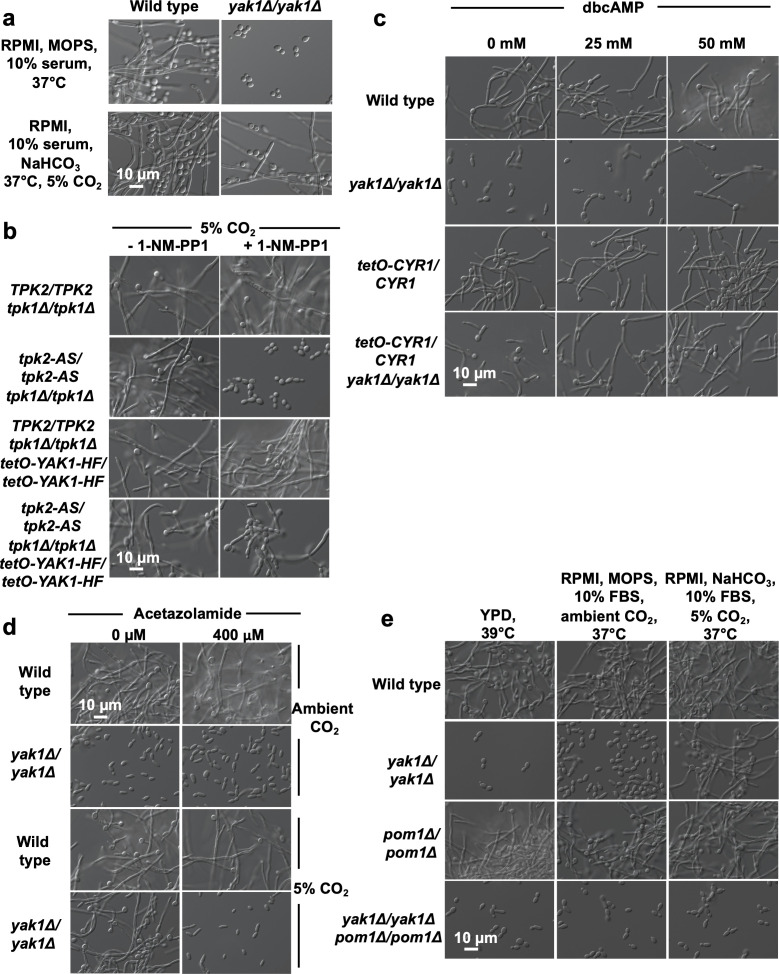
Candida albicans is an opportunistic human fungal pathogen that frequently causes life-threatening infections in immunocompromised individuals. To cause disease, the fungus employs several virulence traits, including its ability to transition between yeast and filamentous states. Previous work identified a role for the kinase Yak1 in regulating C. albicans filamentation. Here, we demonstrate that Yak1 regulates morphogenesis through the canonical cAMP/PKA pathway and that this regulation is environmentally contingent, as host-relevant concentrations of CO2 bypass the requirement of Yak1 for C. albicans morphogenesis. We show a related kinase, Pom1, is important for filamentation in the absence of Yak1 under these host-relevant conditions, as deletion of both genes blocked filamentous growth under all conditions tested. Finally, we demonstrate that Yak1 is required for filamentation in a mouse model of C. albicans dermatitis using genetic and pharmacological approaches. Overall, our results expand our understanding of how Yak1 regulates an important virulence trait in C. albicans.
Keywords: Candida albicans; DYRK; Ras1/PKA; Yak1 kinase; dermatitis; fungal pathogen; morphogenesis.
Conflict of interest statement
L.E.C. and L.W. are co-founders and shareholders in Bright Angel Therapeutics, a platform company for the development of antifungal therapeutics. L.E.C. is a Science Advisor for Kapoose Creek, a company seeking to harness the therapeutic potential of fungi.
Figures






References
-
- Riquelme M, Aguirre J, Bartnicki-García S, Braus GH, Feldbrügge M, Fleig U, Hansberg W, Herrera-Estrella A, Kämper J, Kück U, Mouriño-Pérez RR, Takeshita N, Fischer R. 2018. Fungal morphogenesis, from the polarized growth of hyphae to complex reproduction and infection structures. Microbiol Mol Biol Rev 82:e00068-17. doi: 10.1128/MMBR.00068-17 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- R01 AI127375/AI/NIAID NIH HHS/United States
- FDN-154288/Gouvernement du Canada | Canadian Institutes of Health Research (IRSC)
- R01AI127375/HHS | National Institutes of Health (NIH)
- 189663/W. Garfield Weston Foundation (The W. Garfield Weston Foundation)
- 301617/W. Garfield Weston Foundation (The W. Garfield Weston Foundation)
LinkOut - more resources
Full Text Sources
Medical
