

Longitudinal single-cell transcriptional dynamics throughout neurodegeneration in SCA1
- PMID: 38016472
- PMCID: PMC10922326
- DOI: 10.1016/j.neuron.2023.10.039
Longitudinal single-cell transcriptional dynamics throughout neurodegeneration in SCA1
Abstract
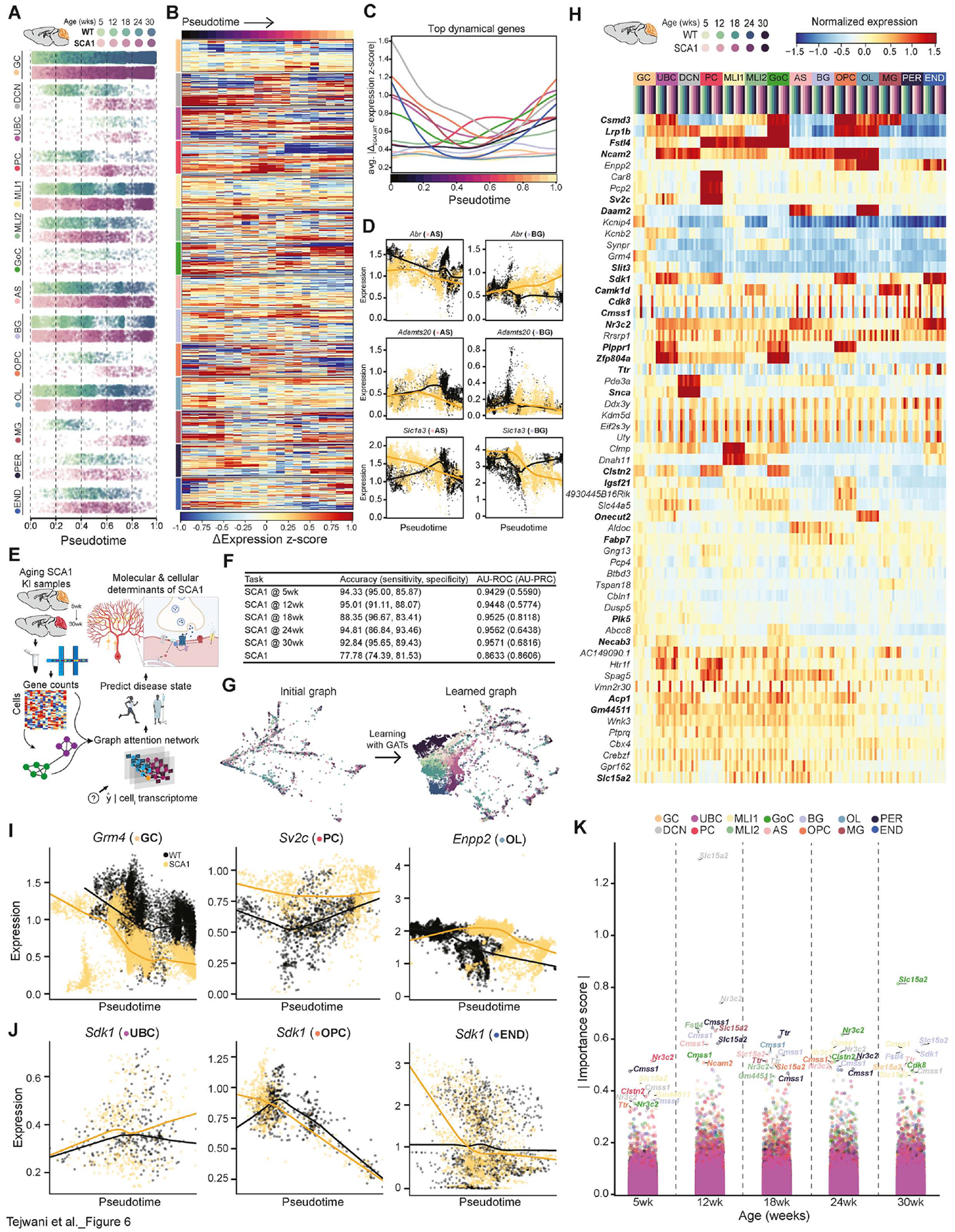
Neurodegeneration is a protracted process involving progressive changes in myriad cell types that ultimately results in the death of vulnerable neuronal populations. To dissect how individual cell types within a heterogeneous tissue contribute to the pathogenesis and progression of a neurodegenerative disorder, we performed longitudinal single-nucleus RNA sequencing of mouse and human spinocerebellar ataxia type 1 (SCA1) cerebellar tissue, establishing continuous dynamic trajectories of each cell population. Importantly, we defined the precise transcriptional changes that precede loss of Purkinje cells and, for the first time, identified robust early transcriptional dysregulation in unipolar brush cells and oligodendroglia. Finally, we applied a deep learning method to predict disease state accurately and identified specific features that enable accurate distinction of wild-type and SCA1 cells. Together, this work reveals new roles for diverse cerebellar cell types in SCA1 and provides a generalizable analysis framework for studying neurodegeneration.
Keywords: Purkinje cell; SCA1; ataxin-1; machine learning; neurodegeneration; oligodendrocyte; oligodendrocyte progenitor cell; single-nucleus RNA sequencing; spinocerebellar ataxia type 1; unipolar brush cells.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.W. is an inventor on a patent applied for by Harvard University related to MERFISH and a consultant and shareholder of Translura, Inc.
Figures





References
-
- Watase K, Weeber EJ, Xu B, Antalffy B, Yuva-Paylor L, Hashimoto K, Kano M, Atkinson R, Sun Y, Armstrong DL, et al. (2002). A Long CAG Repeat in the Mouse Sca1 Locus Replicates SCA1 Features and Reveals the Impact of Protein Solubility on Selective Neurodegeneration. Neuron 34, 905–919. 10.1016/s0896-6273(02)00733-x. - DOI - PubMed
MeSH terms
Substances
Grants and funding
- P30 AG053760/AG/NIA NIH HHS/United States
- R21 MH119803/MH/NIMH NIH HHS/United States
- R01 NS083706/NS/NINDS NIH HHS/United States
- T32 NS041228/NS/NINDS NIH HHS/United States
- R01 AG076154/AG/NIA NIH HHS/United States
- R01 HG011245/HG/NHGRI NIH HHS/United States
- R01 NS027699/NS/NINDS NIH HHS/United States
- R01 AG066447/AG/NIA NIH HHS/United States
- R01 AG074609/AG/NIA NIH HHS/United States
- T32 NS007224/NS/NINDS NIH HHS/United States
- R01 NS088321/NS/NINDS NIH HHS/United States
- R01 NS085054/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
