

Analysis of the indispensable RAD51 cofactor BRCA2 in Naganishia liquefaciens, a Basidiomycota yeast
- PMID: 38016757
- PMCID: PMC10684384
- DOI: 10.26508/lsa.202302342
Analysis of the indispensable RAD51 cofactor BRCA2 in Naganishia liquefaciens, a Basidiomycota yeast
Abstract
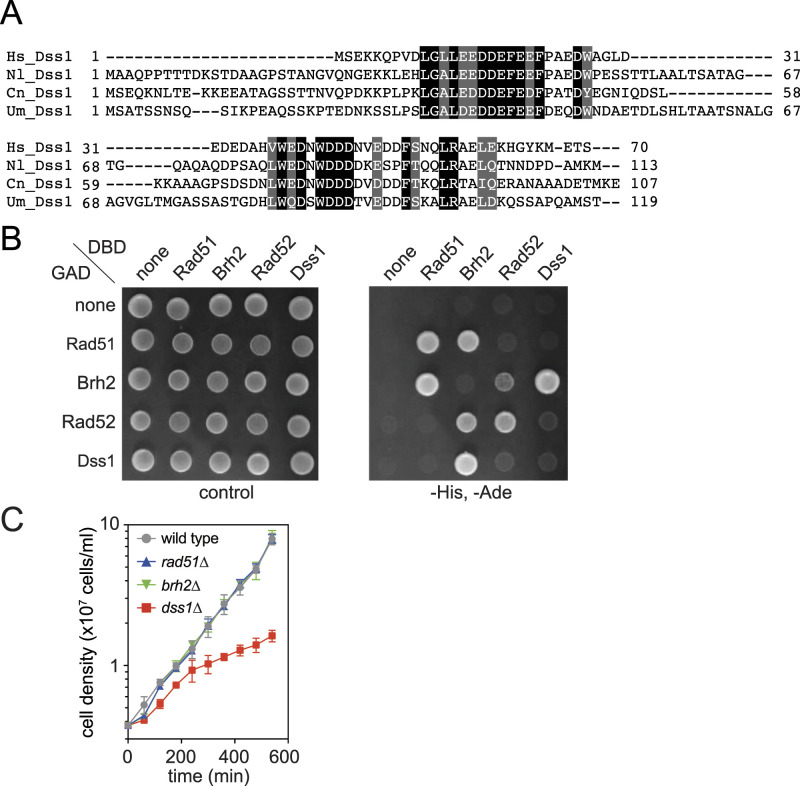
The BRCA2 tumor suppressor plays a critical role in homologous recombination by regulating RAD51, the eukaryotic homologous recombinase. We identified the BRCA2 homolog in a Basidiomycota yeast, Naganishia liquefaciens BRCA2 homologs are found in many Basidiomycota species but not in Ascomycota species. Naganishia BRCA2 (Brh2, for BRCA2 homolog) is about one-third the size of human BRCA2. Brh2 carries three potential BRC repeats with two oligonucleotide/oligosaccharide-binding domains. The homolog of DSS1, a small acidic protein serving as an essential partner of BRCA2 was also identified. The yeast two-hybrid assay shows the interaction of Brh2 with both Rad51 and Dss1. Unlike human BRCA2, Brh2 is not required for normal cell growth, whereas loss of Dss1 results in slow growth. The loss of Brh2 caused pronounced sensitivity to UV and ionizing radiation, and their HR ability, as assayed by gene-targeting efficiency, is compromised. These phenotypes are indistinguishable from those of the rad51 mutant, and the rad51 brh2 double mutant. Naganishia Brh2 is likely the BRCA2 ortholog that functions as an indispensable auxiliary factor for Rad51.
© 2023 Palihati et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures





Similar articles
-
Brh2-Dss1 interplay enables properly controlled recombination in Ustilago maydis.Mol Cell Biol. 2005 Apr;25(7):2547-57. doi: 10.1128/MCB.25.7.2547-2557.2005. Mol Cell Biol. 2005. PMID: 15767662 Free PMC article.
-
Approaches to Understanding the Mediator Function of Brh2 in Ustilago maydis.Methods Enzymol. 2018;600:513-525. doi: 10.1016/bs.mie.2017.11.019. Epub 2018 Feb 1. Methods Enzymol. 2018. PMID: 29458772
-
DNA-binding Domain within the Brh2 N Terminus Is the Primary Interaction Site for Association with DNA.J Biol Chem. 2009 Mar 27;284(13):8265-73. doi: 10.1074/jbc.M809226200. Epub 2009 Feb 1. J Biol Chem. 2009. PMID: 19182269 Free PMC article.
-
The homologous recombination system of Ustilago maydis.Fungal Genet Biol. 2008 Aug;45 Suppl 1(Suppl 1):S31-9. doi: 10.1016/j.fgb.2008.04.006. Epub 2008 May 23. Fungal Genet Biol. 2008. PMID: 18502156 Free PMC article. Review.
-
BRCA2: a universal recombinase regulator.Oncogene. 2007 Dec 10;26(56):7720-30. doi: 10.1038/sj.onc.1210870. Oncogene. 2007. PMID: 18066084 Review.
References
-
- Abe F, Miura T, Nagahama T, Inoue A, Usami R, Horikoshi K (2001) Isolation of a highly copper-tolerant yeast, Cryptococcus sp., from the Japan Trench and the induction of superoxide dismutase activity by Cu2+. Biotechnol Lett 23: 2027–2034. 10.1023/A:1013739232093 - DOI
-
- Antoniou A, Pharoah PDP, Narod S, Risch HA, Eyfjord JE, Hopper JL, Loman N, Olsson H, Johannsson O, Borg Å, et al. (2003) Average risks of breast and ovarian cancer associated with BRCA1 or BRCA2 mutations detected in case series unselected for family history: A combined analysis of 22 studies. Am J Hum Genet 72: 1117–1130. 10.1086/375033 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous