The genetic relationships between brain structure and schizophrenia
- PMID: 38016951
- PMCID: PMC10684873
- DOI: 10.1038/s41467-023-43567-7
The genetic relationships between brain structure and schizophrenia
Abstract
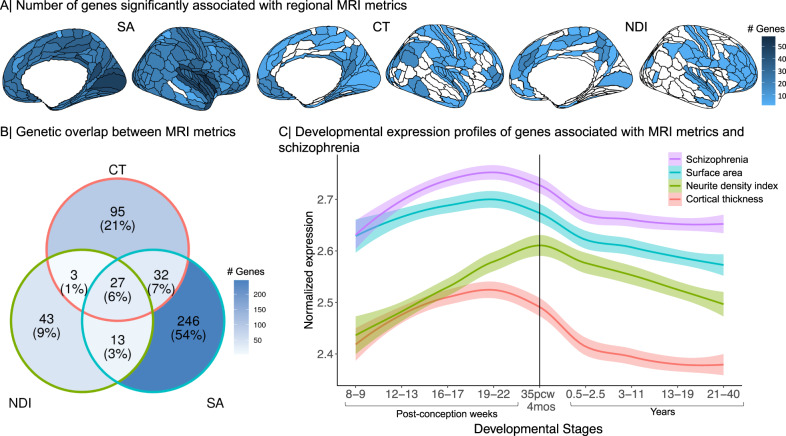
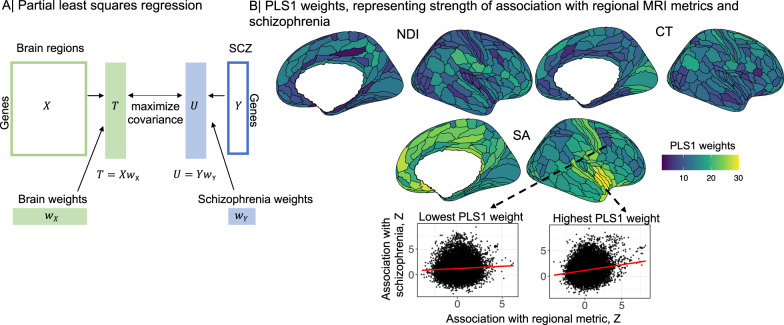
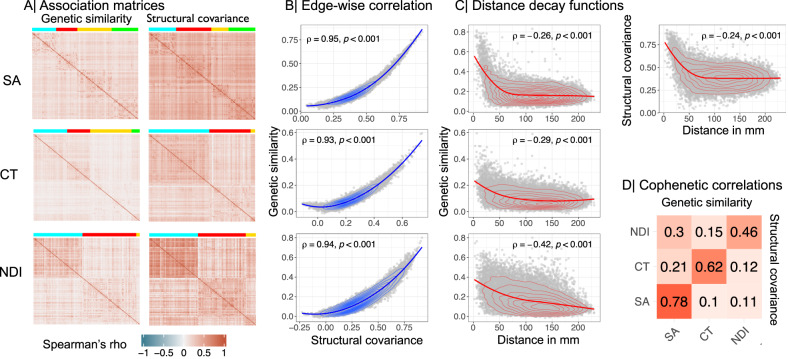
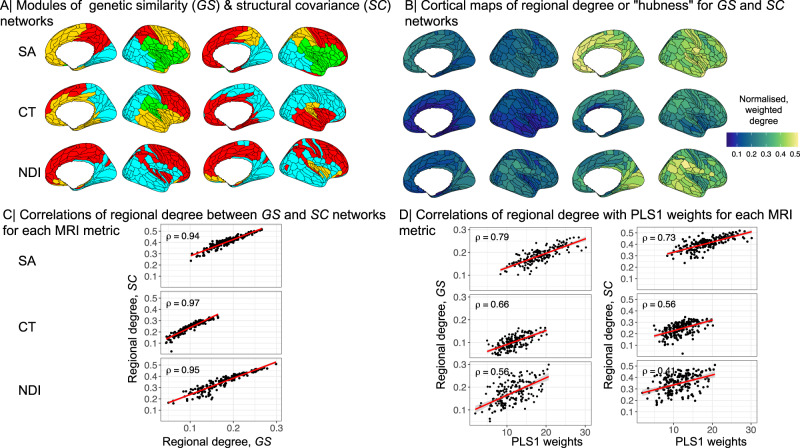
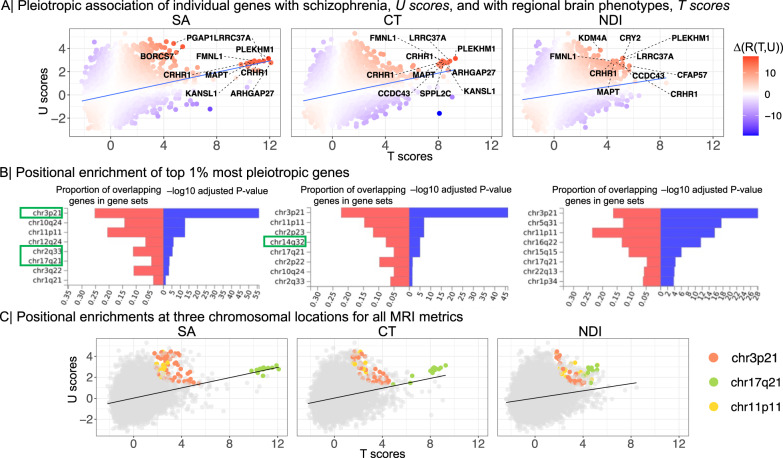
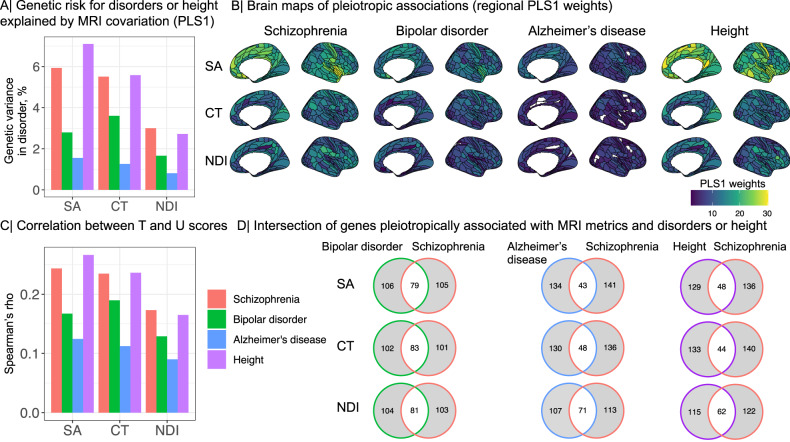
Genetic risks for schizophrenia are theoretically mediated by genetic effects on brain structure but it has been unclear which genes are associated with both schizophrenia and cortical phenotypes. We accessed genome-wide association studies (GWAS) of schizophrenia (N = 69,369 cases; 236,642 controls), and of three magnetic resonance imaging (MRI) metrics (surface area, cortical thickness, neurite density index) measured at 180 cortical areas (N = 36,843, UK Biobank). Using Hi-C-coupled MAGMA, 61 genes were significantly associated with both schizophrenia and one or more MRI metrics. Whole genome analysis with partial least squares demonstrated significant genetic covariation between schizophrenia and area or thickness of most cortical regions. Genetic similarity between cortical areas was strongly coupled to their phenotypic covariance, and genetic covariation between schizophrenia and brain phenotypes was strongest in the hubs of structural covariance networks. Pleiotropically associated genes were enriched for neurodevelopmental processes and positionally concentrated in chromosomes 3p21, 17q21 and 11p11. Mendelian randomization analysis indicated that genetically determined variation in a posterior cingulate cortical area could be causal for schizophrenia. Parallel analyses of GWAS on bipolar disorder, Alzheimer's disease and height showed that pleiotropic association with MRI metrics was stronger for schizophrenia compared to other disorders.
© 2023. The Author(s).
Conflict of interest statement
E.T.B. has consulted for GlaxoSmithKline, SR One, Boehringer Ingelheim, Sosei Heptares, and Monument Therapeutics. R.A.I.B. and E.T.B. are directors of and hold stock in CentileBio. All other authors declare no conflicts of interest.
Figures