

Rhythmic profile of memory T and B-cells along childhood and adolescence
- PMID: 38017254
- PMCID: PMC10684863
- DOI: 10.1038/s41598-023-48115-3
Rhythmic profile of memory T and B-cells along childhood and adolescence
Abstract
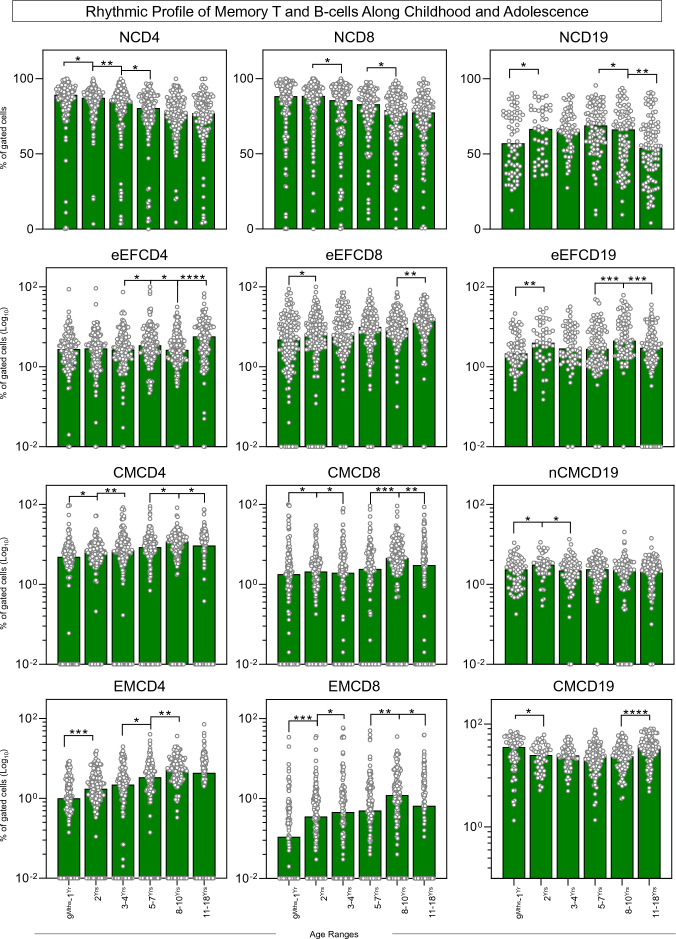
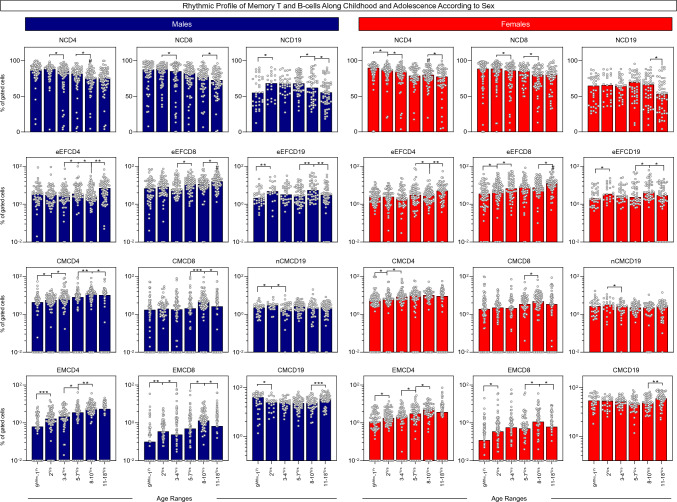
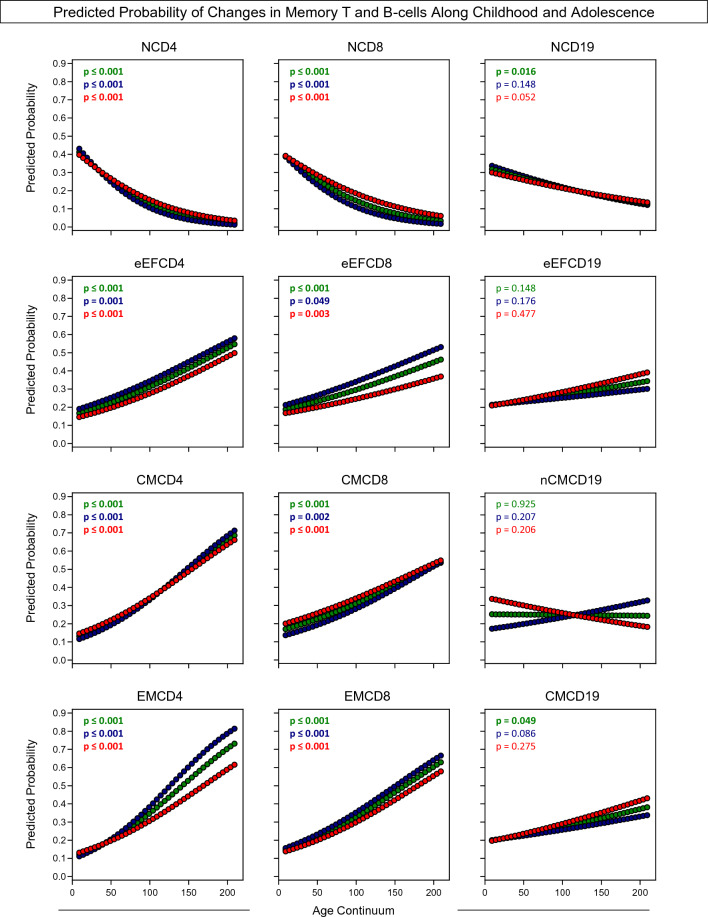
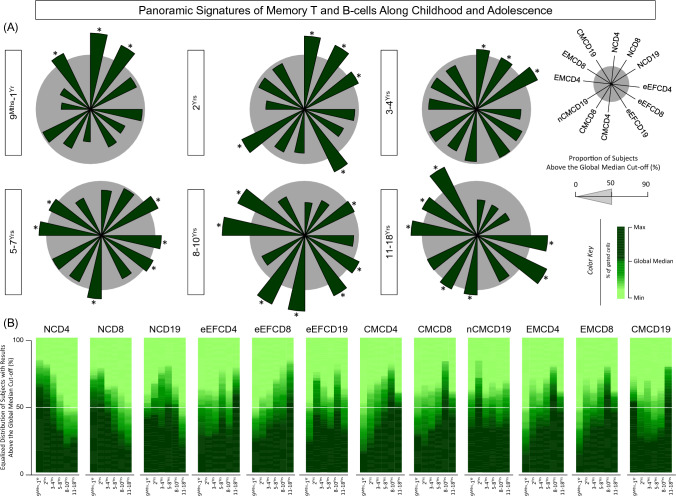
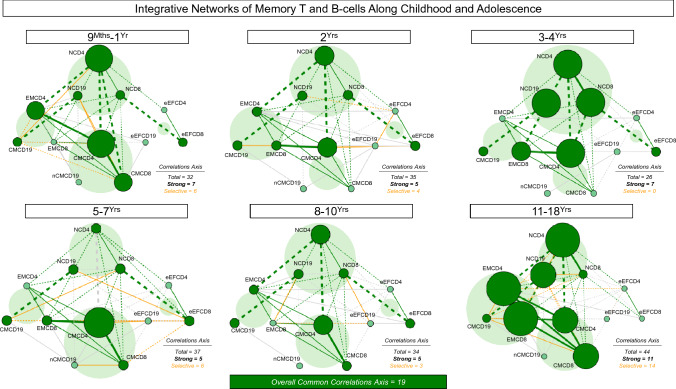
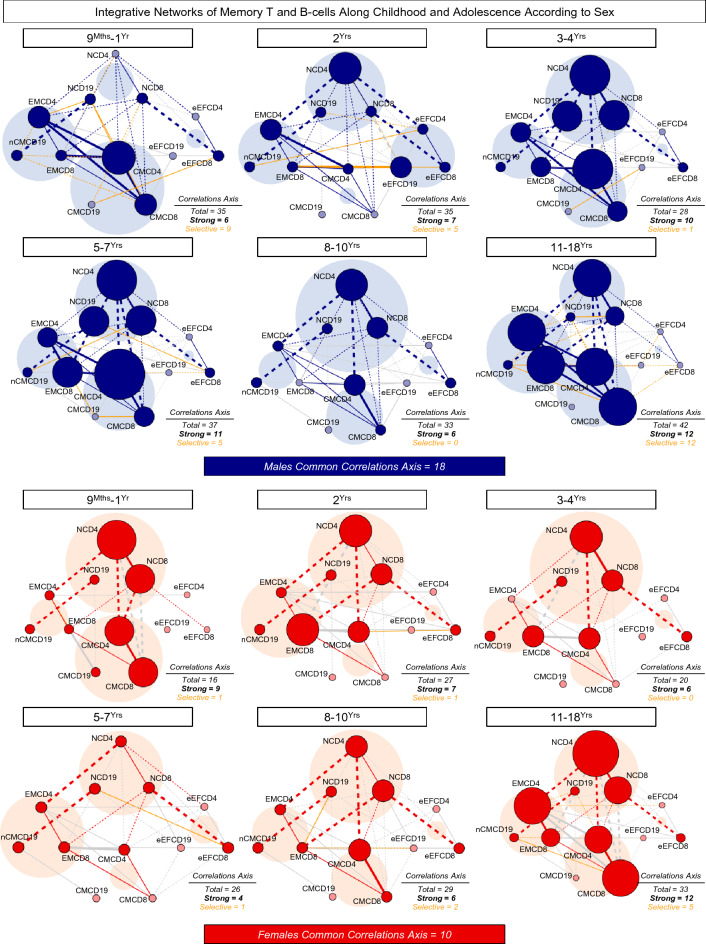
Immunobiography describes the life-long effects of exogenous or endogenous stimuli on remodeling of immune cell biology, including the development of memory T and B-cells. The present study aimed at investigating the rhythms of changes in phenotypic features of memory T and B-cells along childhood and adolescence. A descriptive-observational investigation was conducted including 812 healthy volunteers, clustered into six consecutive age groups (9Mths-1Yr; 2Yrs; 3-4Yrs; 5-7Yrs; 8-10Yrs; 11-18Yrs). Immunophenotypic analysis of memory T-cell (CD4+ and CD8+) and B-cell subsets were performed by flow cytometry. The results pointed out that memory-related biomarkers of T and B-cells displayed a bimodal profile along healthy childhood and adolescence, regardless of sex. The first stage of changes occurs around 2Yrs, with predominance of naive cells, while the second and more prominent wave occurs around the age 8-10Yrs, with the prevalence of memory phenotypes. The neighborhood connectivity profile analysis demonstrated that the number of correlations reaches a peak at 11-18Yrs and lower values along the childhood. Males presented higher and conserved number of correlations when compared to females. Altogether, our results provide new insights into immunobiography and a better understanding of interactions among the cellular subsets studied here during childhood and adolescence.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
