

Flipped C-Terminal Ends of APOA1 Promote ABCA1-Dependent Cholesterol Efflux by Small HDLs
- PMID: 38018436
- PMCID: PMC10913861
- DOI: 10.1161/CIRCULATIONAHA.123.065959
Flipped C-Terminal Ends of APOA1 Promote ABCA1-Dependent Cholesterol Efflux by Small HDLs
Abstract
Background: Cholesterol efflux capacity (CEC) predicts cardiovascular disease independently of high-density lipoprotein (HDL) cholesterol levels. Isolated small HDL particles are potent promoters of macrophage CEC by the ABCA1 (ATP-binding cassette transporter A1) pathway, but the underlying mechanisms are unclear.
Methods: We used model system studies of reconstituted HDL and plasma from control and lecithin-cholesterol acyltransferase (LCAT)-deficient subjects to investigate the relationships among the sizes of HDL particles, the structure of APOA1 (apolipoprotein A1) in the different particles, and the CECs of plasma and isolated HDLs.
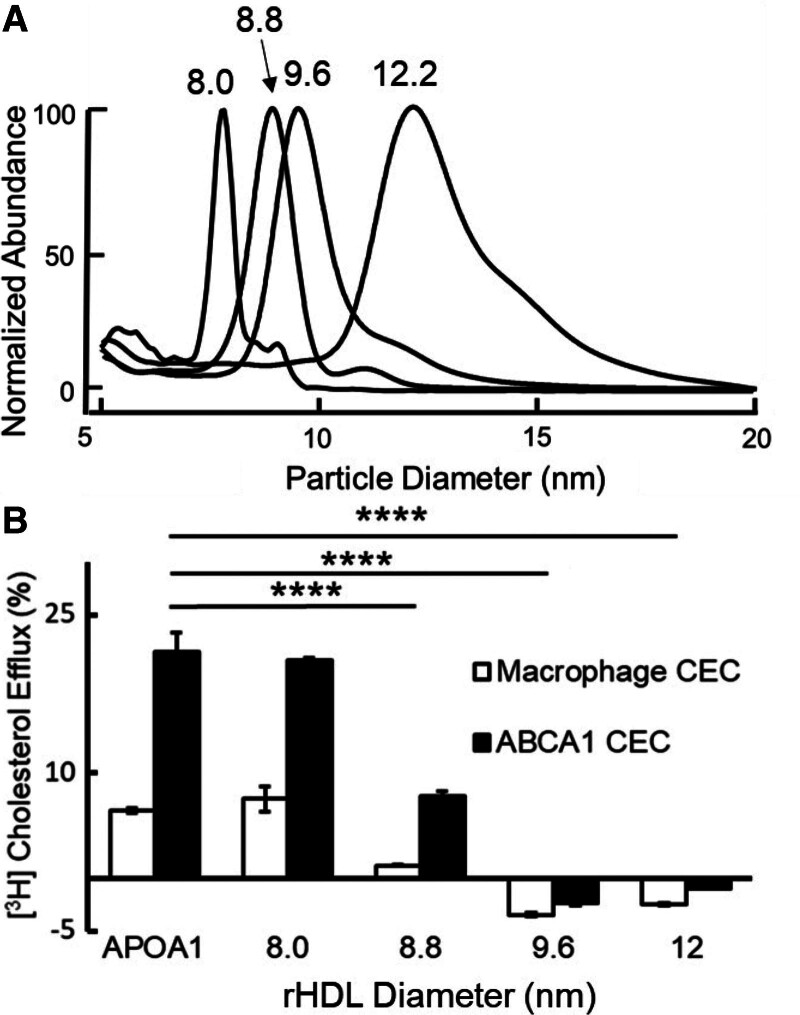
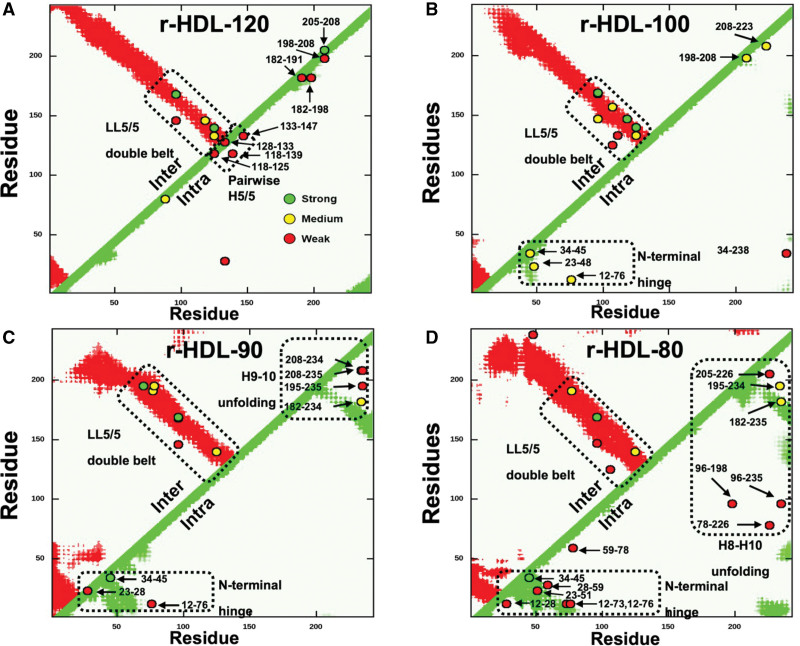
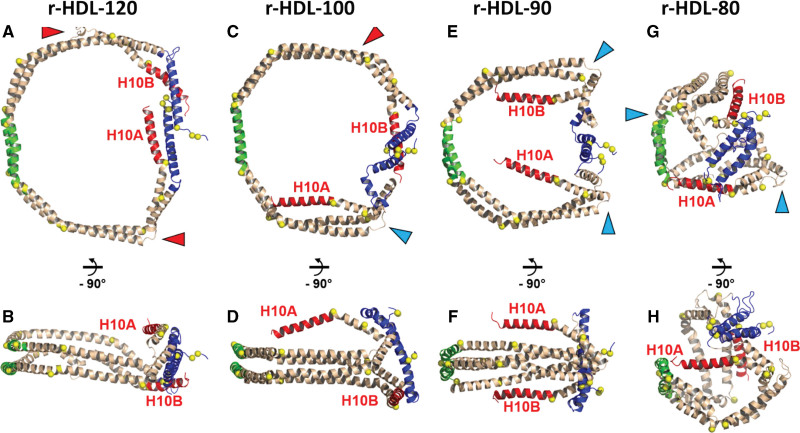
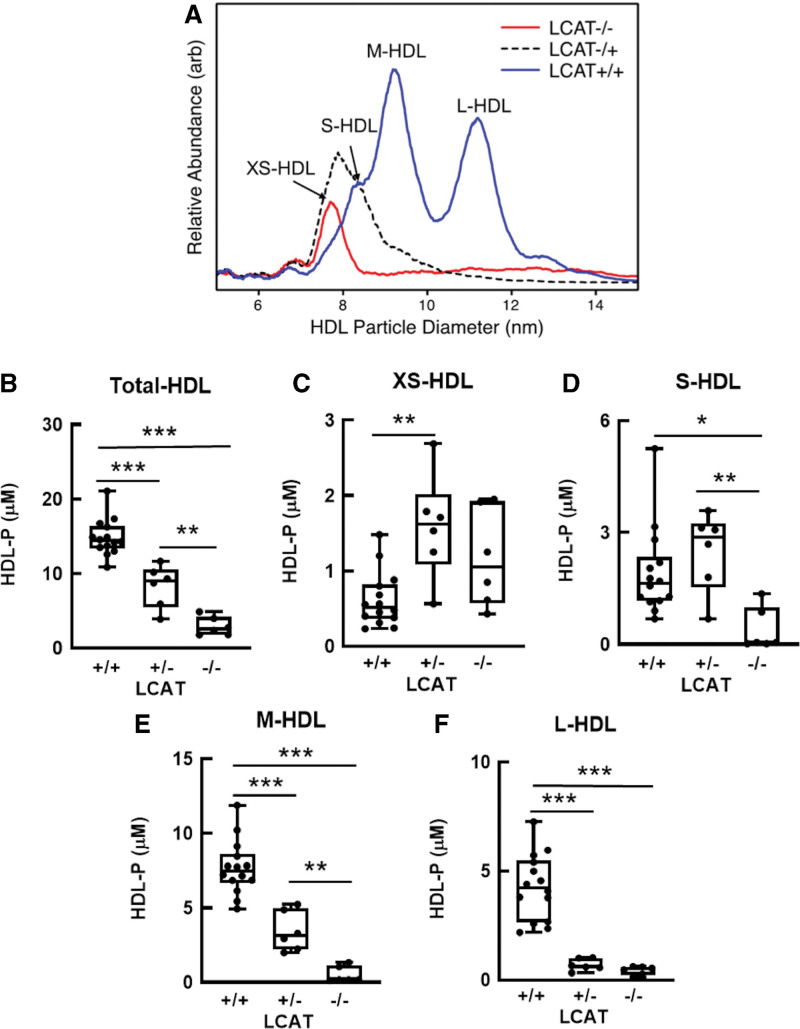
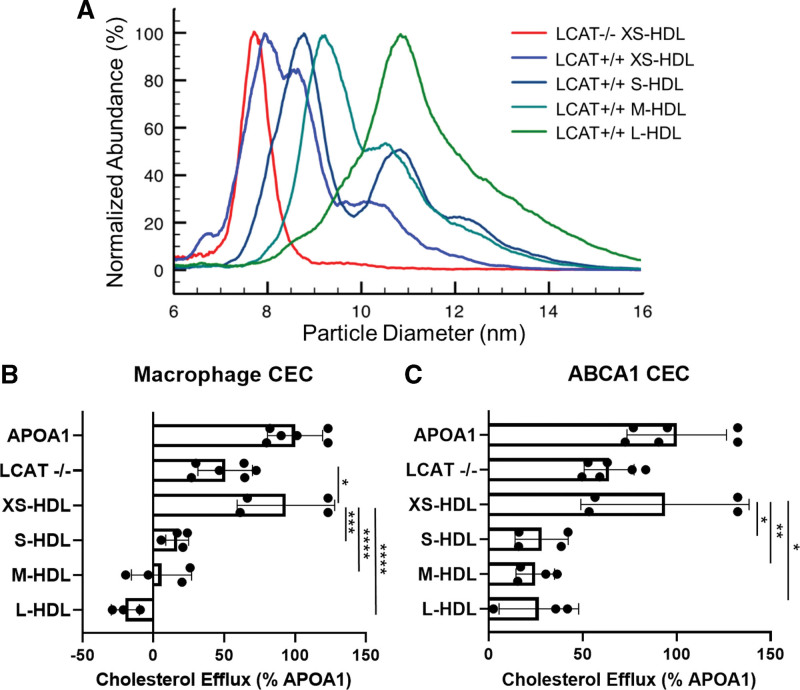
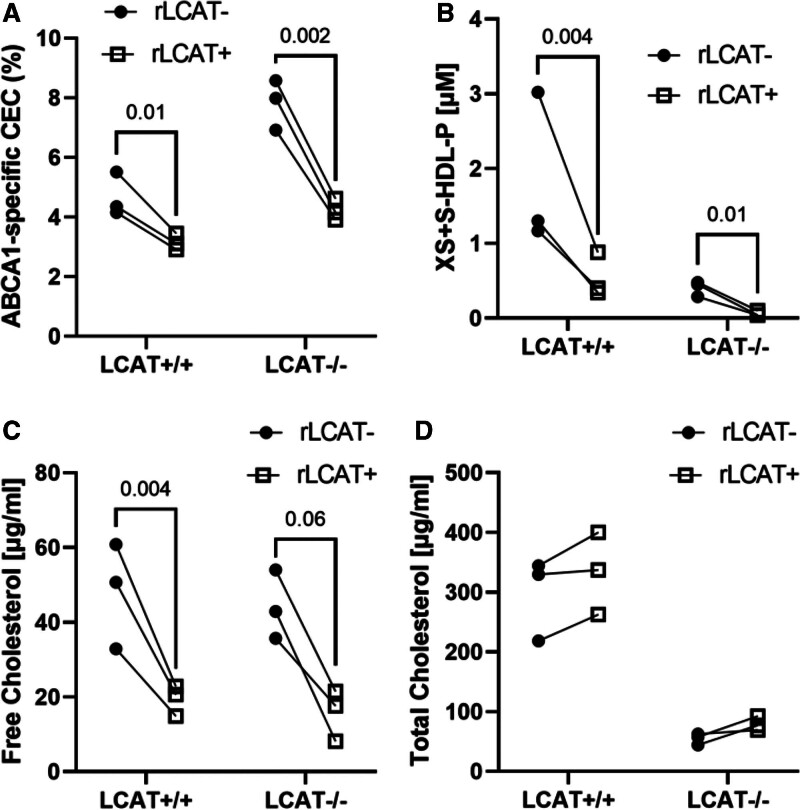
Results: We quantified macrophage and ABCA1 CEC of 4 distinct sizes of reconstituted HDL. CEC increased as particle size decreased. Tandem mass spectrometric analysis of chemically cross-linked peptides and molecular dynamics simulations of APOA1, the major protein of HDL, indicated that the mobility of C-terminus of that protein was markedly higher and flipped off the surface in the smallest particles. To explore the physiological relevance of the model system studies, we isolated HDL from LCAT-deficient subjects, whose small HDLs (like reconstituted HDLs) are discoidal and composed of APOA1, cholesterol, and phospholipid. Despite their very low plasma levels of HDL particles, these subjects had normal CEC. In both the LCAT-deficient subjects and control subjects, the CEC of isolated extra-small HDL (a mixture of extra-small and small HDL by calibrated ion mobility analysis) was 3- to 5-fold greater than that of the larger sizes of isolated HDL. Incubating LCAT-deficient plasma and control plasma with human LCAT converted extra-small and small HDL particles into larger particles, and it markedly inhibited CEC.
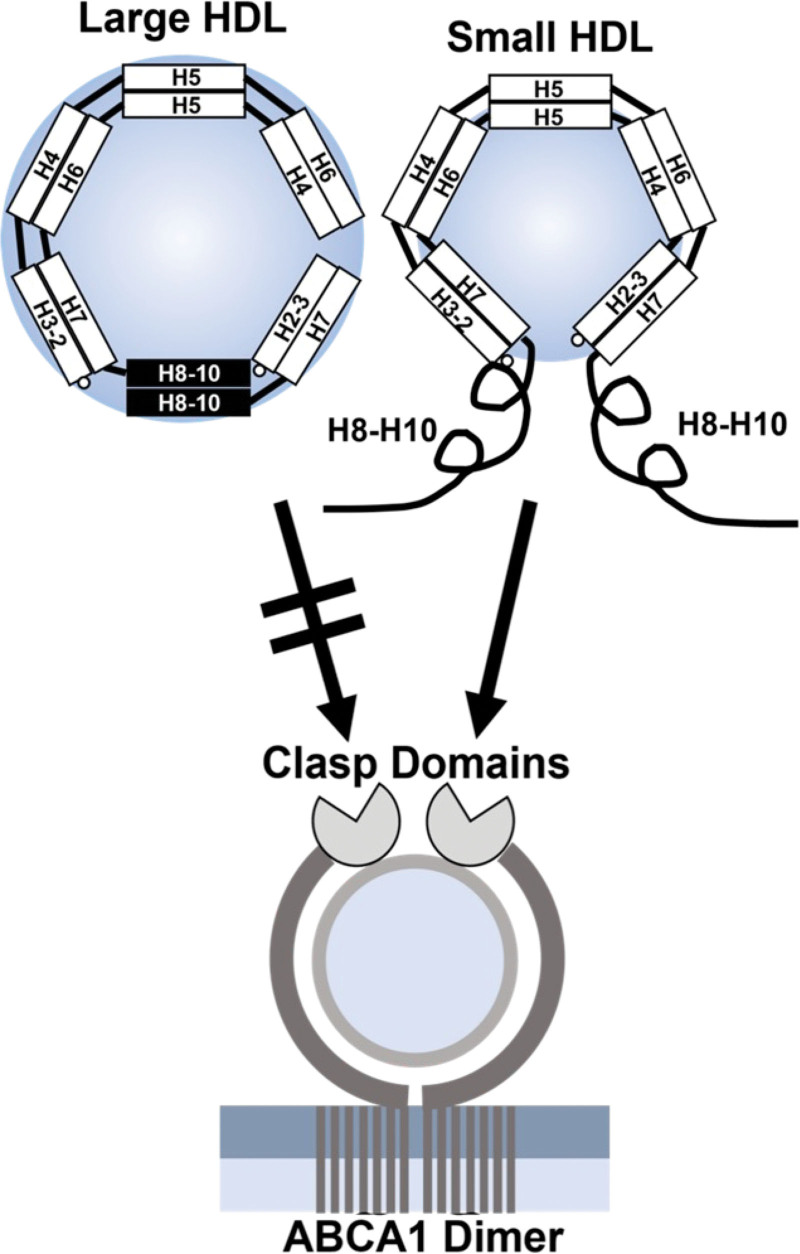
Conclusions: We present a mechanism for the enhanced CEC of small HDLs. In smaller particles, the C-termini of the 2 antiparallel molecules of APOA1 are "flipped" off the lipid surface of HDL. This extended conformation allows them to engage with ABCA1. In contrast, the C-termini of larger HDLs are unable to interact productively with ABCA1 because they form a helical bundle that strongly adheres to the lipid on the particle. Enhanced CEC, as seen with the smaller particles, predicts decreased cardiovascular disease risk. Thus, extra-small and small HDLs may be key mediators and indicators of the cardioprotective effects of HDL.
Keywords: ABCA1; atherosclerosis; cholesterol efflux capacity; computational biology; lipids and cholesterol.
Conflict of interest statement
Figures
Update of
-
Flipped C-Terminal Ends of APOA1 Promote ABCA1-dependent Cholesterol Efflux by Small HDLs.medRxiv [Preprint]. 2023 Nov 4:2023.11.03.23297986. doi: 10.1101/2023.11.03.23297986. medRxiv. 2023. Update in: Circulation. 2024 Mar 5;149(10):774-787. doi: 10.1161/CIRCULATIONAHA.123.065959. PMID: 37961344 Free PMC article. Updated. Preprint.
References
-
- Gordon DJ, Rifkind BM. High-density lipoprotein--the clinical implications of recent studies. N Engl J Med. 1989;321:1311–1316. doi: 10.1056/NEJM198911093211907 - PubMed
-
- Rader DJ, Hovingh GK. HDL and cardiovascular disease. Lancet. 2014;384:618–625. doi: 10.1016/S0140-6736(14)61217-4 - PubMed
-
- Rader DJ, Tall AR. The not-so-simple HDL story: is it time to revise the HDL cholesterol hypothesis? Nat Med. 2012;18:1344–1346. doi: 10.1038/nm.2937 - PubMed
-
- Heinecke J. HDL and cardiovascular-disease risk--time for a new approach? N Engl J Med. 2011;364:170–171. doi: 10.1056/NEJMe1012520 - PubMed
