

An injury-responsive mmp14b enhancer is required for heart regeneration
- PMID: 38019918
- PMCID: PMC10686572
- DOI: 10.1126/sciadv.adh5313
An injury-responsive mmp14b enhancer is required for heart regeneration
Abstract
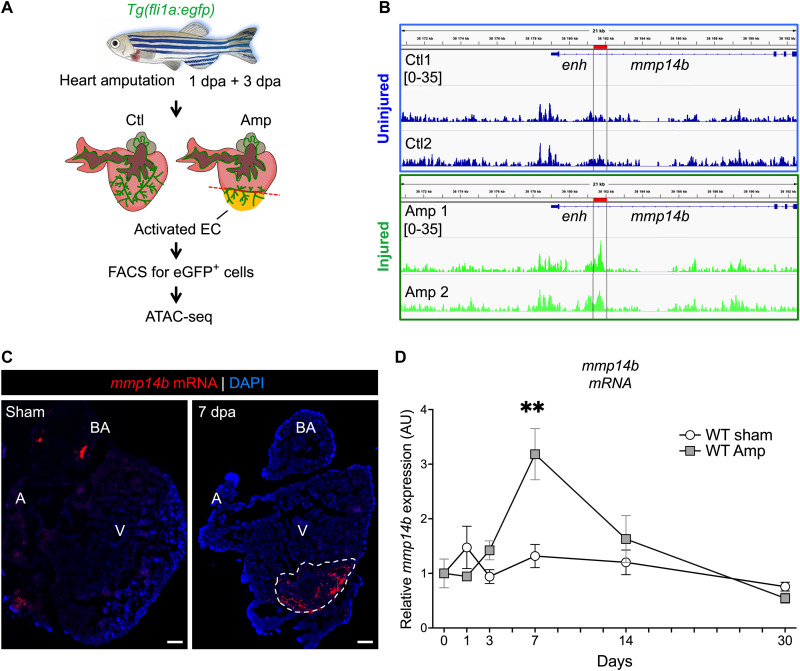
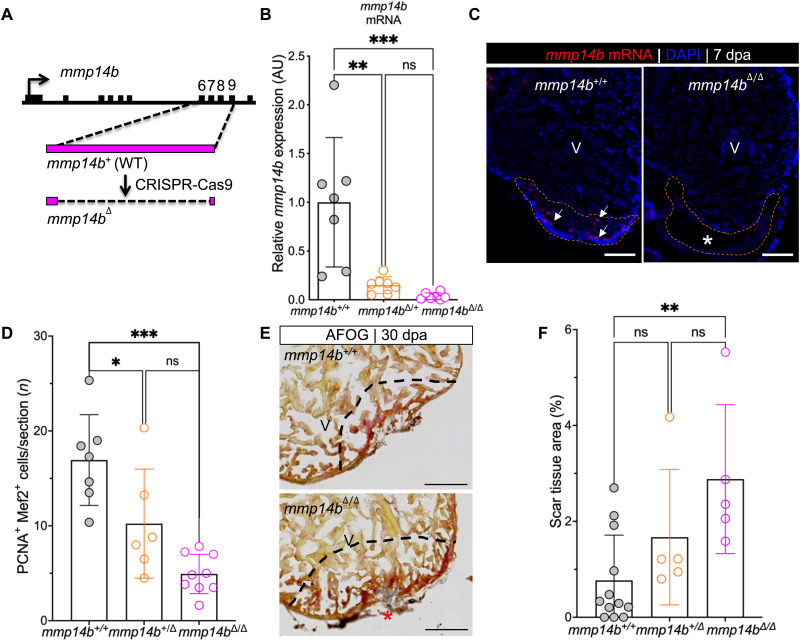
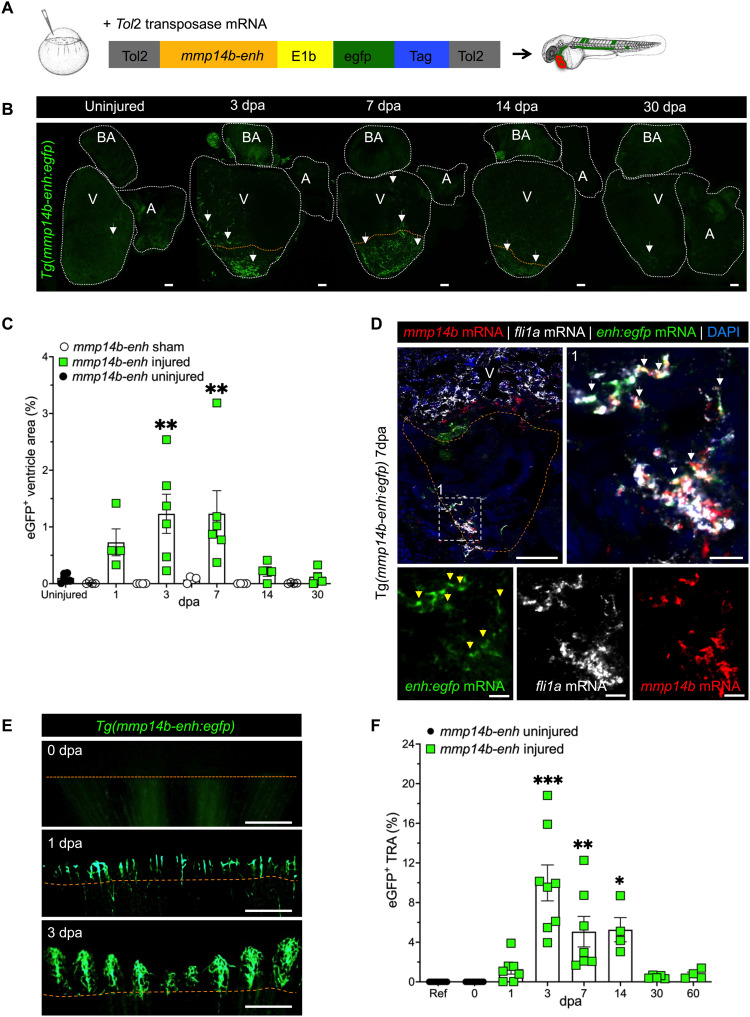
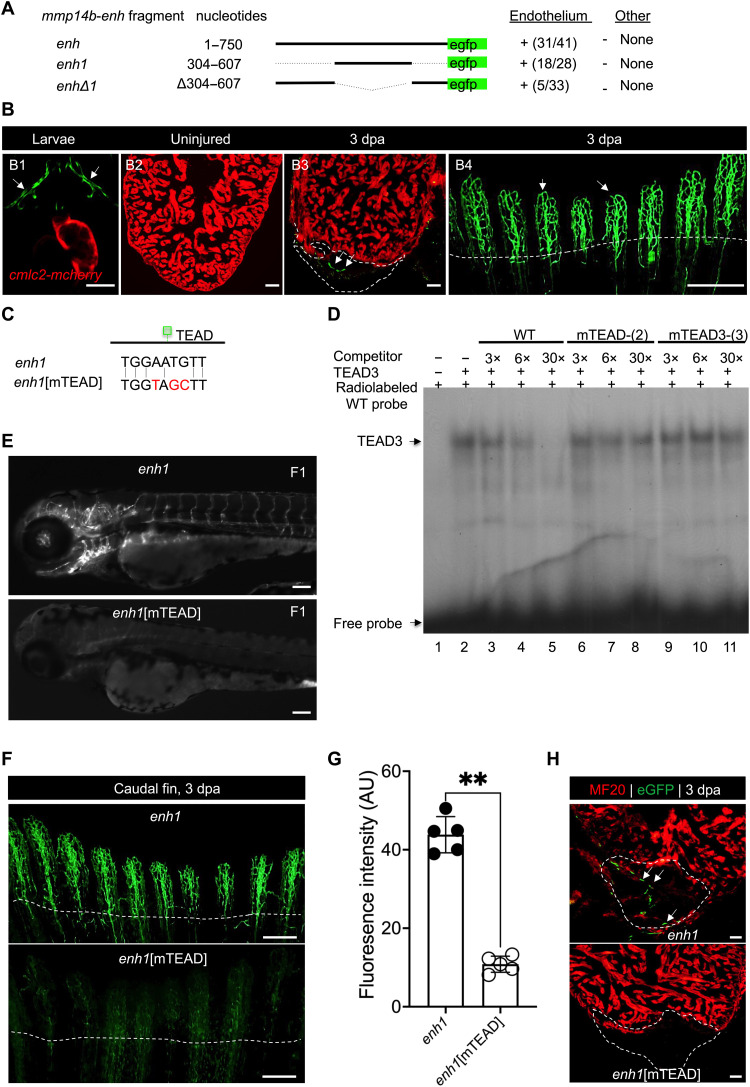
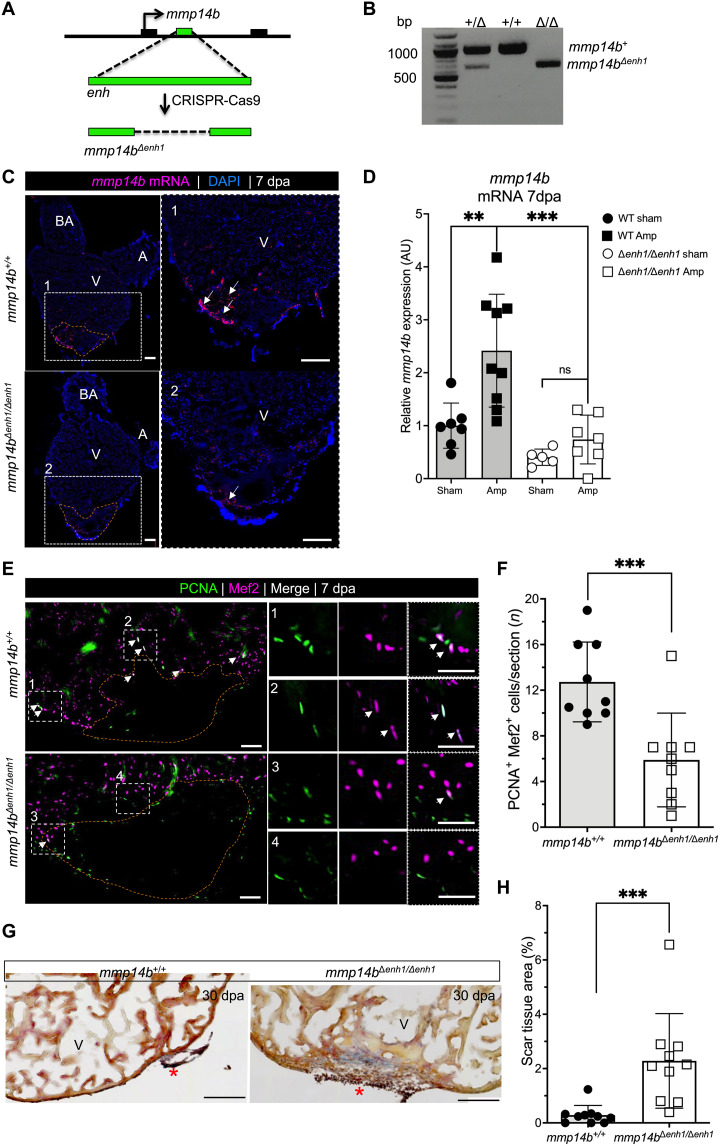
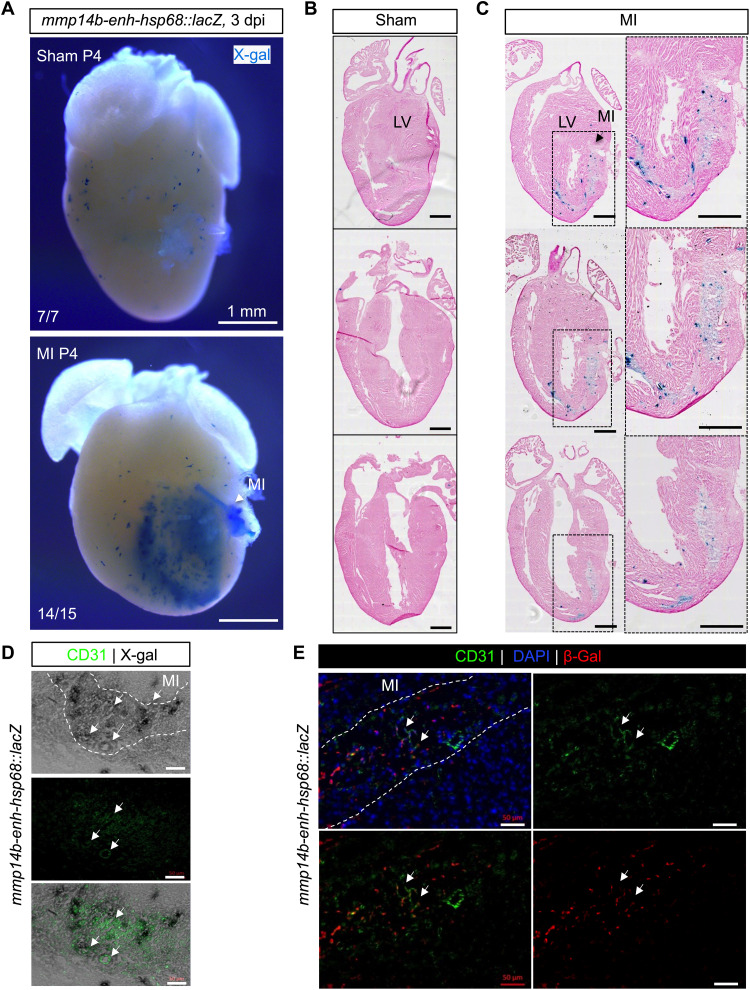
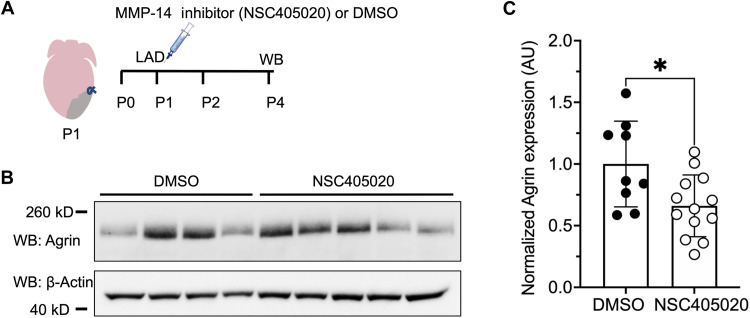
Mammals have limited capacity for heart regeneration, whereas zebrafish have extraordinary regeneration abilities. During zebrafish heart regeneration, endothelial cells promote cardiomyocyte cell cycle reentry and myocardial repair, but the mechanisms responsible for promoting an injury microenvironment conducive to regeneration remain incompletely defined. Here, we identify the matrix metalloproteinase Mmp14b as an essential regulator of heart regeneration. We identify a TEAD-dependent mmp14b endothelial enhancer induced by heart injury in zebrafish and mice, and we show that the enhancer is required for regeneration, supporting a role for Hippo signaling upstream of mmp14b. Last, we show that MMP-14 function in mice is important for the accumulation of Agrin, an essential regulator of neonatal mouse heart regeneration. These findings reveal mechanisms for extracellular matrix remodeling that promote heart regeneration.
Figures
References
-
- E. J. Benjamin, S. S. Virani, C. W. Callaway, A. M. Chamberlain, A. R. Chang, S. Cheng, S. E. Chiuve, M. Cushman, F. N. Delling, R. Deo, S. D. de Ferranti, J. F. Ferguson, M. Fornage, C. Gillespie, C. R. Isasi, M. C. Jiménez, L. C. Jordan, S. E. Judd, D. Lackland, J. H. Lichtman, L. Lisabeth, S. Liu, C. T. Longenecker, P. L. Lutsey, J. S. Mackey, D. B. Matchar, K. Matsushita, M. E. Mussolino, K. Nasir, M. O'Flaherty, L. P. Palaniappan, A. Pandey, D. K. Pandey, M. J. Reeves, M. D. Ritchey, C. J. Rodriguez, G. A. Roth, W. D. Rosamond, U. K. A. Sampson, G. M. Satou, S. H. Shah, N. L. Spartano, D. L. Tirschwell, C. W. Tsao, J. H. Voeks, J. Z. Willey, J. T. Wilkins, J. H. Wu, H. M. Alger, S. S. Wong, P. Muntner, Heart Disease and Stroke Statistics-2018 Update: A report from the American Heart Association. Circulation 137, e67–e492 (2018). - PubMed
-
- W. Wang, C. K. Hu, A. Zeng, D. Alegre, D. Hu, K. Gotting, A. Ortega Granillo, Y. Wang, S. Robb, R. Schnittker, S. Zhang, D. Alegre, H. Li, E. Ross, N. Zhang, A. Brunet, A. Sánchez Alvarado, Changes in regeneration-responsive enhancers shape regenerative capacities in vertebrates. Science 369, eaaz3090 (2020). - PMC - PubMed
-
- T. Hoang, J. Wang, P. Boyd, F. Wang, C. Santiago, L. Jiang, S. Yoo, M. Lahne, L. J. Todd, M. Jia, C. Saez, C. Keuthan, I. Palazzo, N. Squires, W. A. Campbell, F. Rajaii, T. Parayil, V. Trinh, D. W. Kim, G. Wang, L. J. Campbell, J. Ash, A. J. Fischer, D. R. Hyde, J. Qian, S. Blackshaw, Gene regulatory networks controlling vertebrate retinal regeneration. Science 370, eabb8598 (2020). - PMC - PubMed
-
- T. Gerber, P. Murawala, D. Knapp, W. Masselink, M. Schuez, S. Hermann, M. Gac-Santel, S. Nowoshilow, J. Kageyama, S. Khattak, J. D. Currie, J. G. Camp, E. M. Tanaka, B. Treutlein, Single-cell analysis uncovers convergence of cell identities during axolotl limb regeneration. Science 362, eaaq0681 (2018). - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 20TPA35500000 - GUO HUANG/AHA/American Heart Association-American Stroke Association/United States
- R01HL160819 (G.N.H.), R01HL138456 (G.N.H.), DK119621 (B.L.B.), AND HL146366 (B.L.B.)./NH/NIH HHS/United States
- R01 DK119621/DK/NIDDK NIH HHS/United States
- R01 HL138456/HL/NHLBI NIH HHS/United States
- R01 HL160819/HL/NHLBI NIH HHS/United States
