

Context-dependent neocentromere activity in synthetic yeast chromosome VIII
- PMID: 38020969
- PMCID: PMC10667555
- DOI: 10.1016/j.xgen.2023.100437
Context-dependent neocentromere activity in synthetic yeast chromosome VIII
Abstract
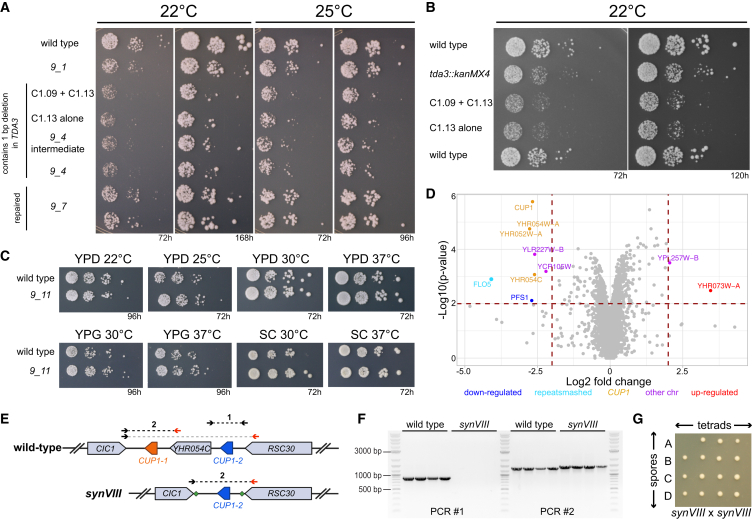
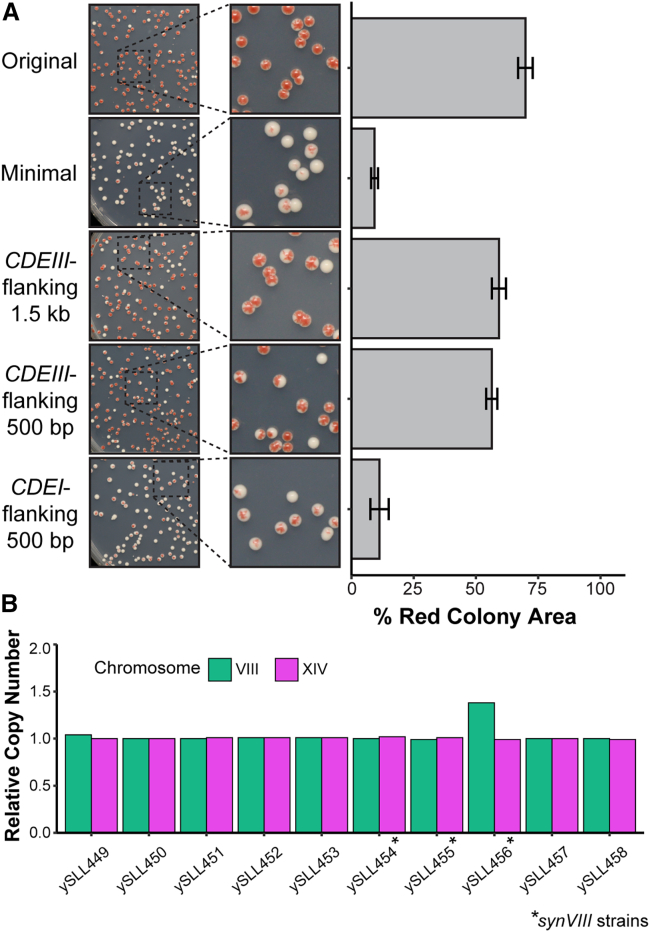
Pioneering advances in genome engineering, and specifically in genome writing, have revolutionized the field of synthetic biology, propelling us toward the creation of synthetic genomes. The Sc2.0 project aims to build the first fully synthetic eukaryotic organism by assembling the genome of Saccharomyces cerevisiae. With the completion of synthetic chromosome VIII (synVIII) described here, this goal is within reach. In addition to writing the yeast genome, we sought to manipulate an essential functional element: the point centromere. By relocating the native centromere sequence to various positions along chromosome VIII, we discovered that the minimal 118-bp CEN8 sequence is insufficient for conferring chromosomal stability at ectopic locations. Expanding the transplanted sequence to include a small segment (∼500 bp) of the CDEIII-proximal pericentromere improved chromosome stability, demonstrating that minimal centromeres display context-dependent functionality.
Keywords: CRISPR; Saccharomyces cerevisiae; aneuploidy; centromere; chromosomal stability; genome engineering; genome rearrangements; pericentromere; synVIII.
© 2023 The Authors.
Conflict of interest statement
J.B. is a founder and director of CDI Labs, Inc., a founder of and consultant to Neochromosome, Inc., a founder, SAB member of, and consultant to ReOpen Diagnostics, LLC, and serves or served on the Scientific Advisory Board of the following: Logomix, Inc., Modern Meadow, Inc., Rome Therapeutics, Inc., Sample6, Inc., Sangamo, Inc., Tessera Therapeutics, Inc., and the Wyss Institute. J.S.B. is a founder of and consultant to Neochromosome. G.S. is a consultant to Neochromosome Inc. and ZenithAI.
Figures




References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
