

Consequences of a telomerase-related fitness defect and chromosome substitution technology in yeast synIX strains
- PMID: 38020974
- PMCID: PMC10667316
- DOI: 10.1016/j.xgen.2023.100419
Consequences of a telomerase-related fitness defect and chromosome substitution technology in yeast synIX strains
Abstract
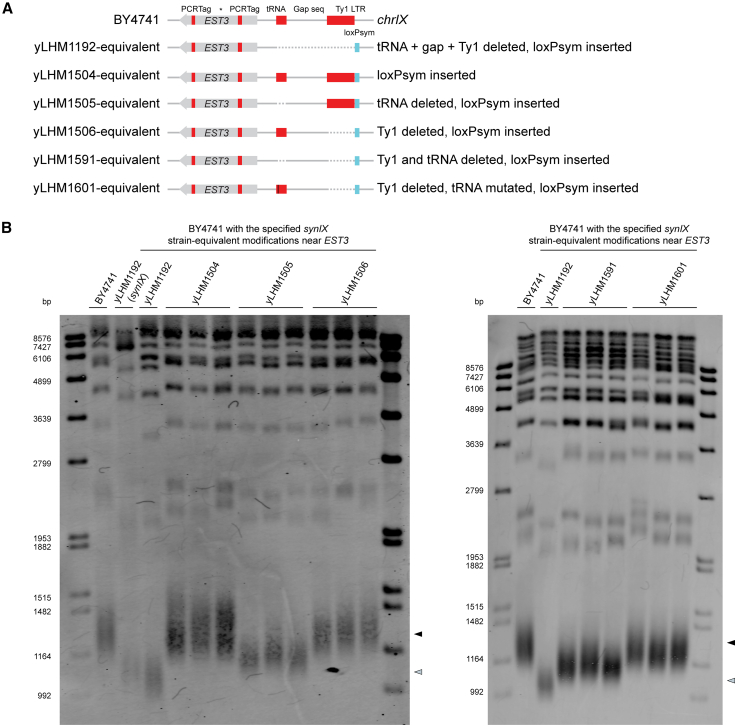
We describe the complete synthesis, assembly, debugging, and characterization of a synthetic 404,963 bp chromosome, synIX (synthetic chromosome IX). Combined chromosome construction methods were used to synthesize and integrate its left arm (synIXL) into a strain containing previously described synIXR. We identified and resolved a bug affecting expression of EST3, a crucial gene for telomerase function, producing a synIX strain with near wild-type fitness. To facilitate future synthetic chromosome consolidation and increase flexibility of chromosome transfer between distinct strains, we combined chromoduction, a method to transfer a whole chromosome between two strains, with conditional centromere destabilization to substitute a chromosome of interest for its native counterpart. Both steps of this chromosome substitution method were efficient. We observed that wild-type II tended to co-transfer with synIX and was co-destabilized with wild-type IX, suggesting a potential gene dosage compensation relationship between these chromosomes.
Keywords: EST3; Saccharomyces cerevisiae; centromere destabilization; chromoduction; chromosome debugging; chromosome substitution; megachunk assembly; synIX; transcriptomics.
© 2023 The Authors.
Conflict of interest statement
J.D.B. is a founder and director of CDI Labs, Inc., a founder of and consultant to Neochromosome, Inc, a founder, SAB member of, and consultant to ReOpen Diagnostics, LLC, and serves or served on the scientific advisory board of the following: Logomix, Inc., Sangamo, Inc., Modern Meadow, Inc., Rome Therapeutics, Inc., Sample6, Inc., Tessera Therapeutics, Inc., and the Wyss Institute. J.S.B. is a founder of Neochromosome, Inc., consultant to Opentrons Labworks, Inc., and advisor to Reflexion Pharmaceuticals, Inc.
Figures







References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
