

Early cellular mechanisms of type I interferon-driven susceptibility to tuberculosis
- PMID: 38029747
- PMCID: PMC10757650
- DOI: 10.1016/j.cell.2023.11.002
Early cellular mechanisms of type I interferon-driven susceptibility to tuberculosis
Abstract
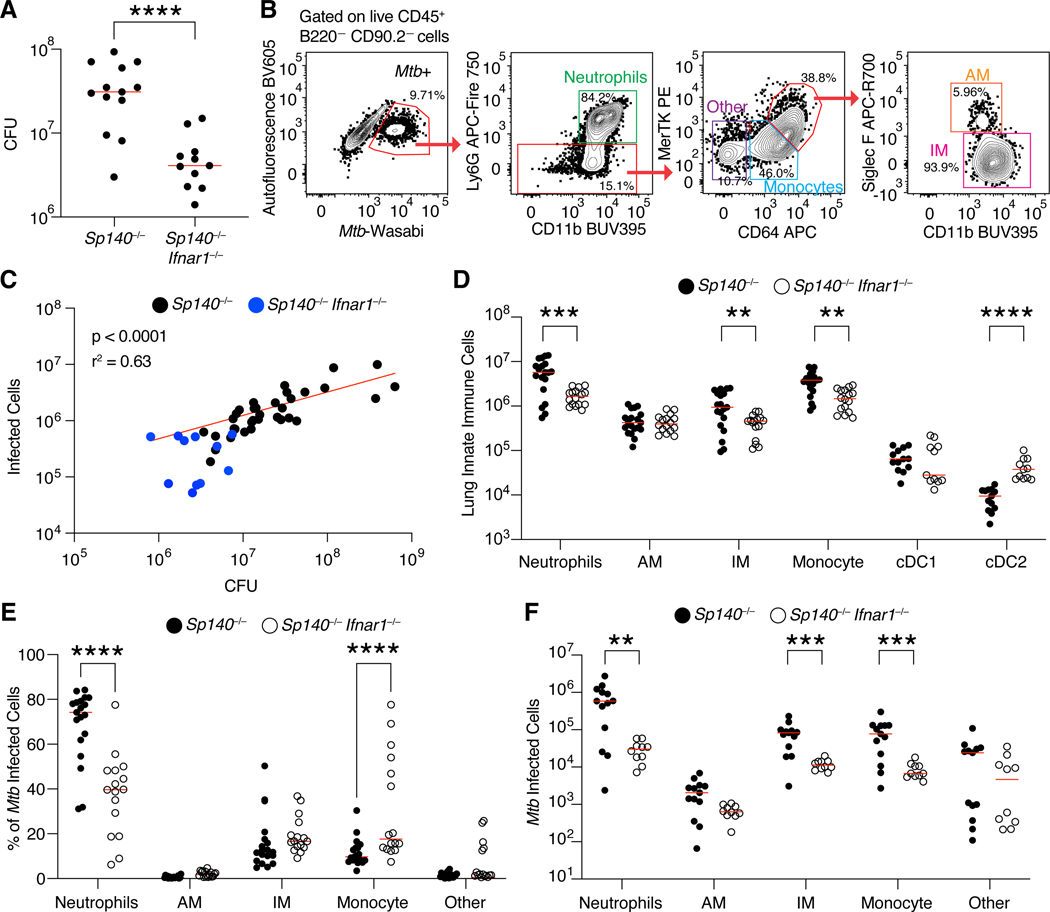
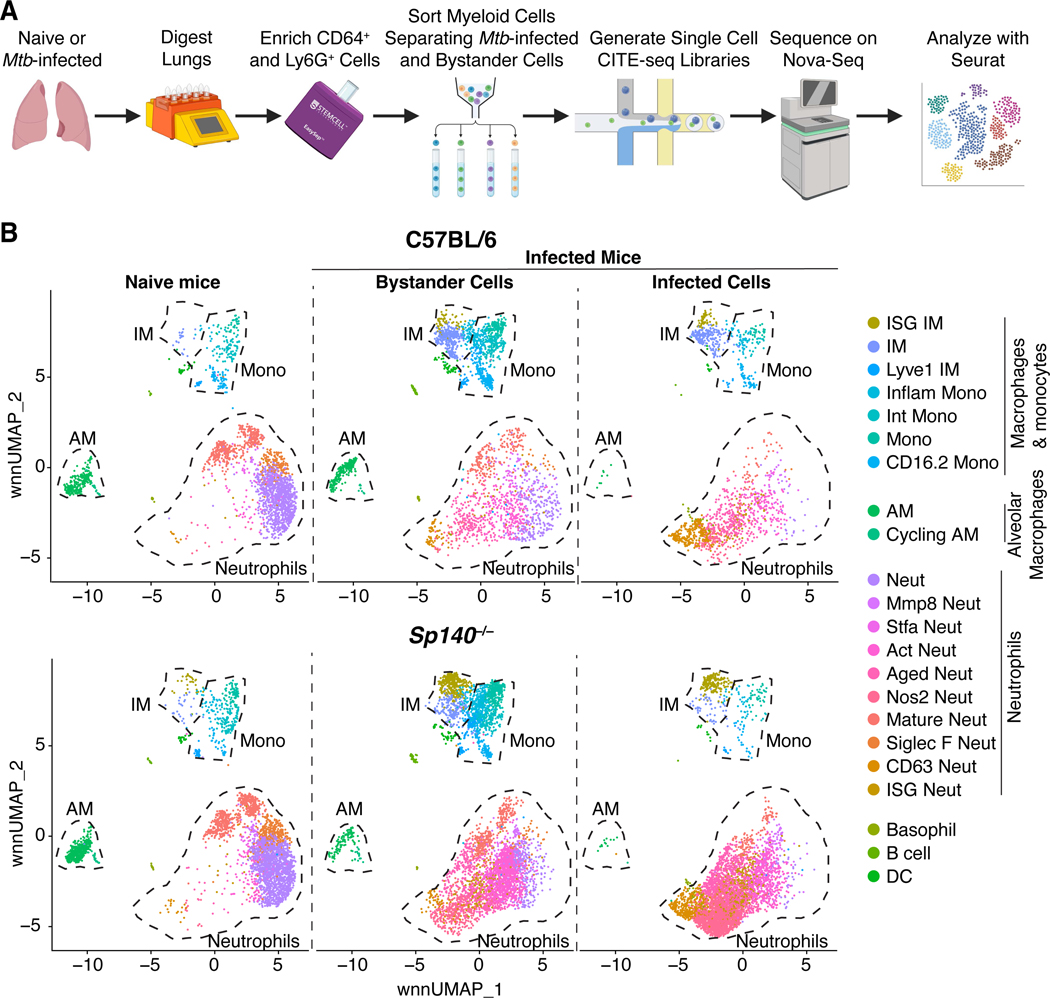
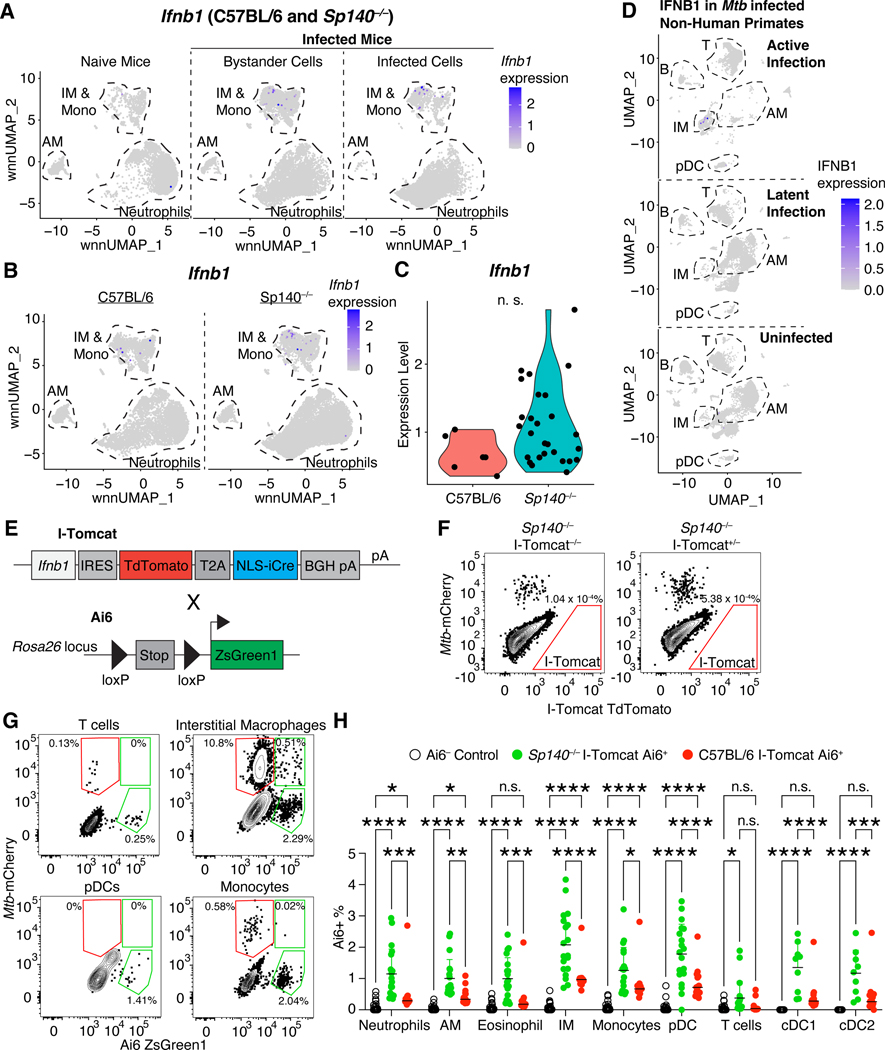
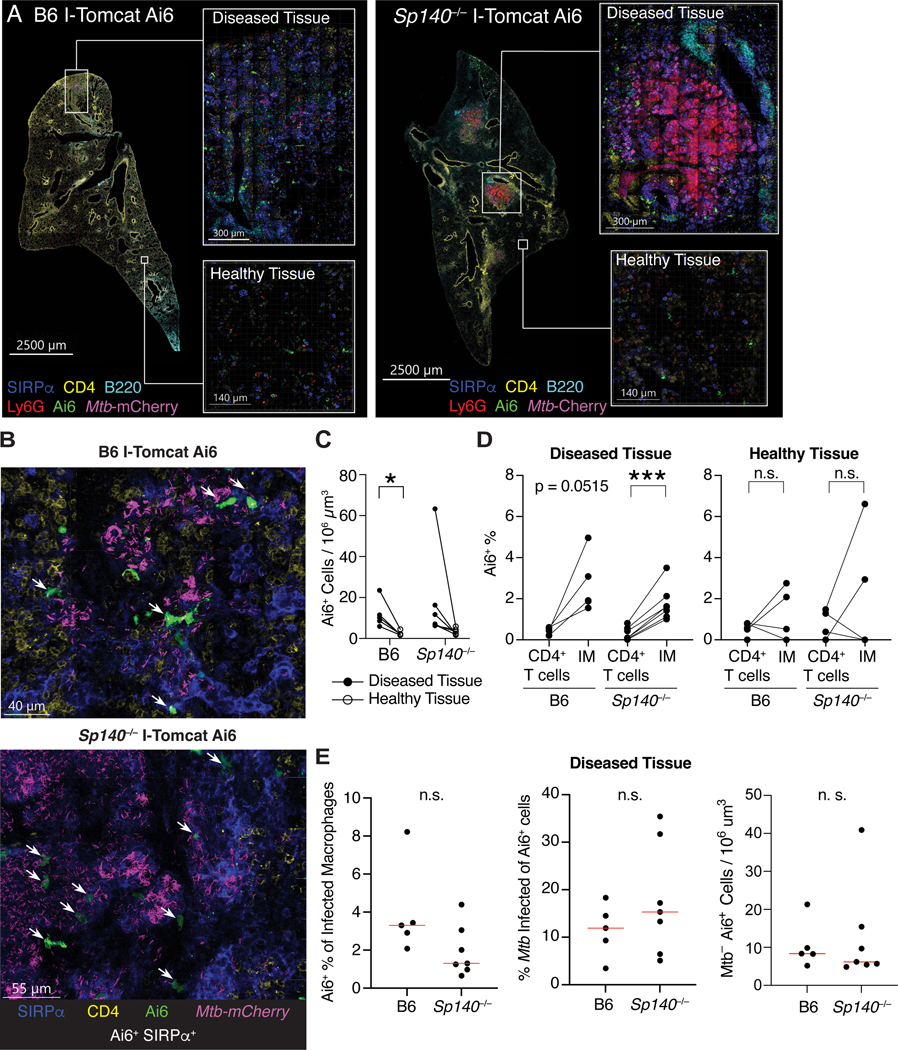
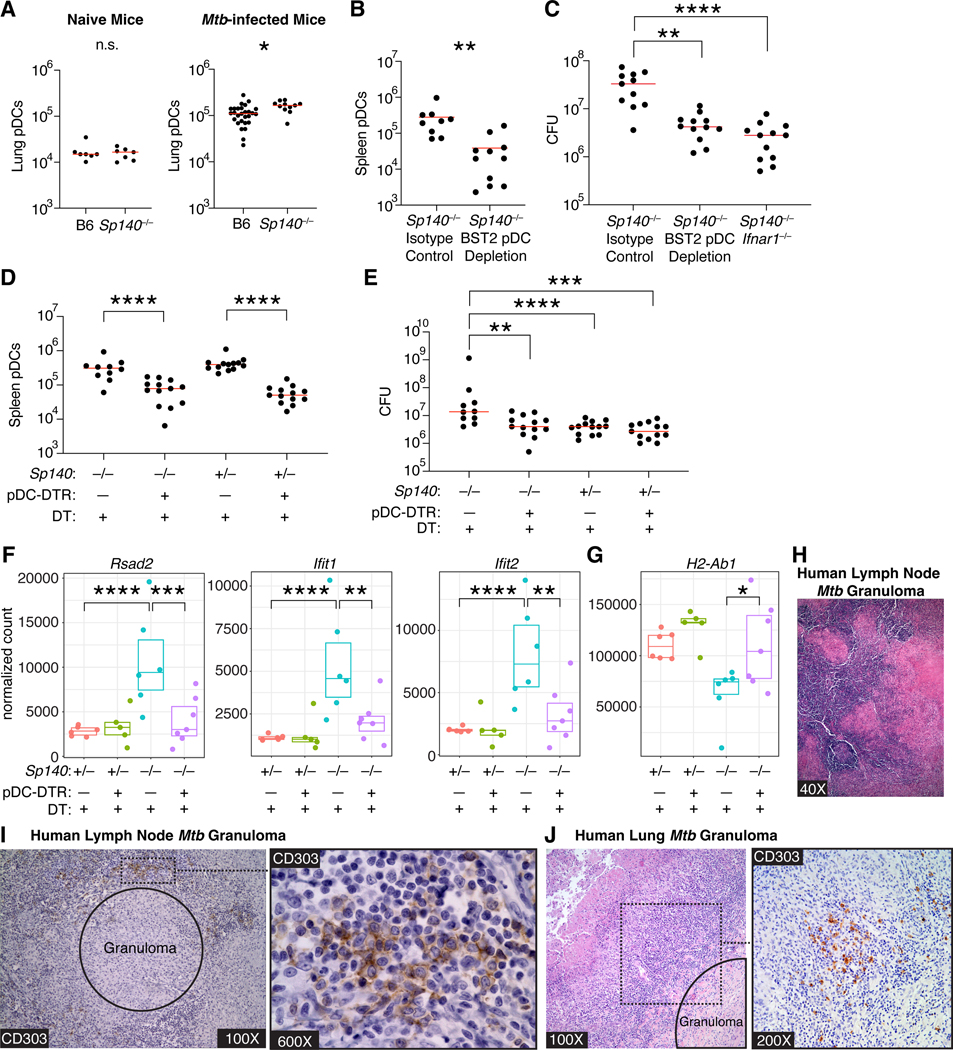
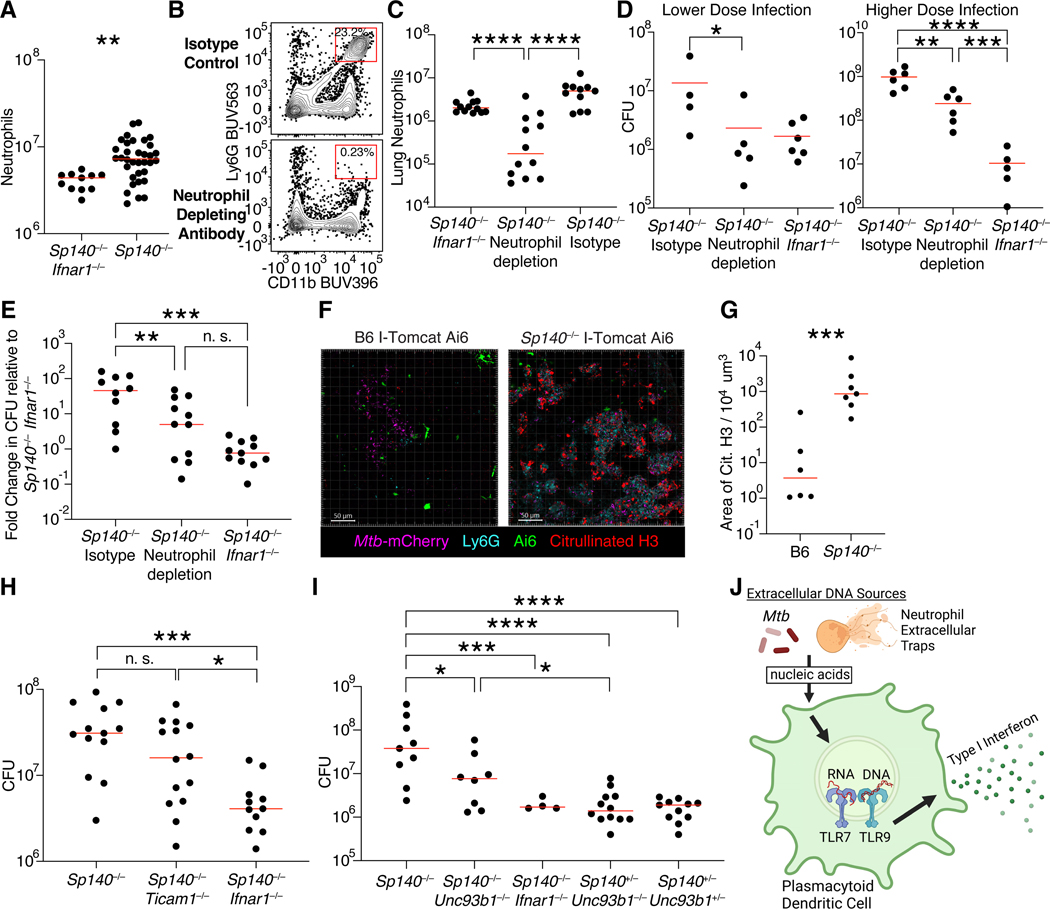
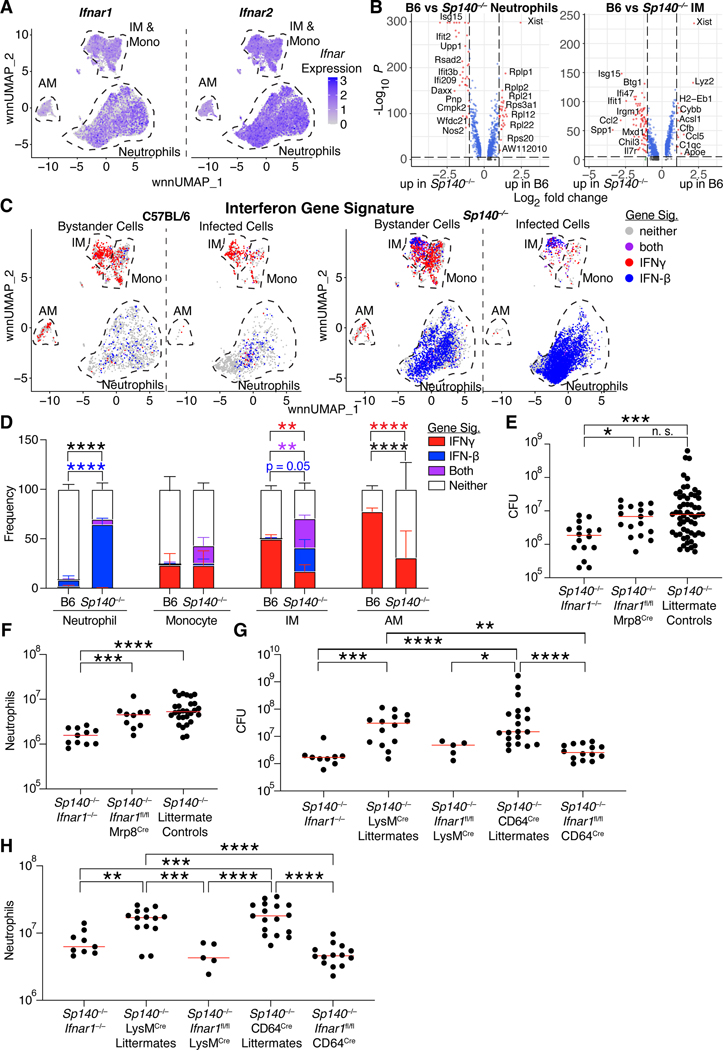
Mycobacterium tuberculosis (Mtb) causes 1.6 million deaths annually. Active tuberculosis correlates with a neutrophil-driven type I interferon (IFN) signature, but the cellular mechanisms underlying tuberculosis pathogenesis remain poorly understood. We found that interstitial macrophages (IMs) and plasmacytoid dendritic cells (pDCs) are dominant producers of type I IFN during Mtb infection in mice and non-human primates, and pDCs localize near human Mtb granulomas. Depletion of pDCs reduces Mtb burdens, implicating pDCs in tuberculosis pathogenesis. During IFN-driven disease, we observe abundant DNA-containing neutrophil extracellular traps (NETs) described to activate pDCs. Cell-type-specific disruption of the type I IFN receptor suggests that IFNs act on IMs to inhibit Mtb control. Single-cell RNA sequencing (scRNA-seq) indicates that type I IFN-responsive cells are defective in their response to IFNγ, a cytokine critical for Mtb control. We propose that pDC-derived type I IFNs act on IMs to permit bacterial replication, driving further neutrophil recruitment and active tuberculosis disease.
Keywords: Mycobacterium tuberculosis; innate immunology; interstitial macrophages; lung; mice; neutrophil extracellular traps; neutrophils; plasmacytoid dendritic cells; type I interferons.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests R.E.V. consults for Ventus Therapeutics, Tempest Therapeutics, and X-biotix Therapeutics.
Figures
References
-
- WHO (2021). Global tuberculosis report 2021. https://www.who.int/publications-detail-redirect/9789240037021.
-
- Katelaris AL, Jackson C, Southern J, Gupta RK, Drobniewski F, Lalvani A, Lipman M, Mangtani P, and Abubakar I. (2020). Effectiveness of BCG Vaccination Against Mycobacterium tuberculosis Infection in Adults: A Cross-sectional Analysis of a UK-Based Cohort. J. Infect. Dis 221, 146–155. 10.1093/infdis/jiz430. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
